КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Транскрипция протяженных межгенных промежутков (нк РНК)
|
|
|
|
Наличие отдаленных энхансеров
Эпигеномное наследование экспрессии некодирующих РНК.
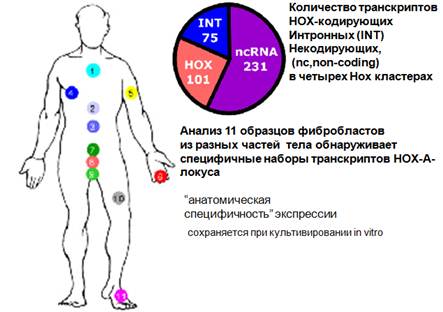
Эпигеномное наследование - это наследование, которое может проявляться и сохраняться после многих делений развития зародыша. В данном случае может сказываться на то, какие гены и кластеры экспрессируются в эпителии у человека в зависимости от того, откуда взяты фибробласты с тела. В этих фибробластах можно исследовать набор транскриптов и видеть, что наборы, взятые с проксимальной части и с дистальной части будут отличаться. Во первых, чем фибробласты интересны, их можно получать в первичной культуре и дальше манипулировать с этими клетками, искать кодирующие и некодирующие транскрипты in vitro. Если анализировать количество транскриптов HOX-кластеров, то основная масса этих транскриптов будет падать на нкРНК (интроны н-р). Когда анализируются транскриптомы из различных частей тела, то выявляется анатомическая специфичность экспрессии, которая очень удобно для исследователя и сохраняется и при клеточном делении in vitro. Специфика, какие гены, когда будут экспрессироваться, сохраняется при развитии зародыша и проявляется уже во взрослом теле.
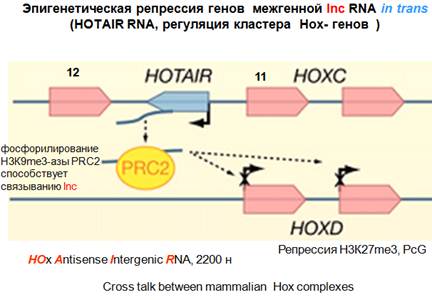
Транс-действие. НОХ-кластер. НОХС и НОХD. Условно антисмысловая транскрипция от транскрипции белок-кодирующих генов. Это интересная нкРНК НOx Antisense Intergenic RNA. Сокращенно HOTAIR. Активно исследуется. Оказалось, что если она экспрессируется, то подавляет экспрессию генов находящихся в другом кластере (транс-действие). Возникает два интересных вопроса. Первый, как HOTAIR может что-то подавлять, какие белковые комплексы она привлекает. На этот вопрос есть ответ. Второй вопрос, как она вообще находит эти места в другой хромосоме, как она передвигается по геному, это вопрос очень интересный, но на него ответа еще нет. Что выяснилось относительно нкРНК, они достаточно длинные, укладываются в определенную структуру, определенные модули и узнаются белками модифицирующими хроматин.
HOTAIR привлекает два разных репрессорных комплекса и координирует модификации гистонов
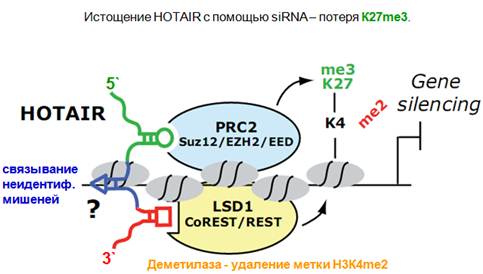
Это все гипотетическая структура, но экспериментально было показано, что с модулем РНК, находящимся на 5’- конце связывается комплекс PRC2, а на 3’- конце связывается комплекс, который определяет деметилирование. Деметелирование специфичное, удаляется метил с четвертого лизина гистона Н3 (Н3К4me2 - метка активного хроматина, которая показывает то, что либо ген транскрибируется, либо то, что он может активно транскрибироваться). Благодаря тому что HOTAIR имеет эти модули, она объединяет за счет взаимодействующих модулей два этих комплекса, которые координированно осуществляют репрессию хроматина. LSD1 собственно удаляет метил с 4-ого лизина, а комплекс PCR2 навешивает метильную метку уже на 27 лизин. Т.о. координировано осуществляется сайленсиг гена. Важен смысл существования такого гибкого скелета РНК, на который можно определенным образом воздействовать. Все эти вещи можно делать в культурах клеток. Истощая HOTAIR с помощью специальной siРНК, получаем уничтожение той метки, которая имеет место при экспрессии этой нкРНК.
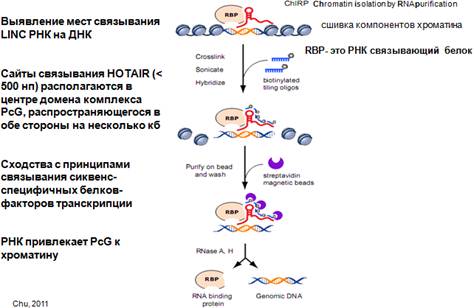
Какая существует методика, попытка выяснить, каким образом нкРНК узнает определенные участки ДНК, куда она может перемещаться. Смотрим на картинку: видим нкРНК, хроматин, РНК-связывающий белок, эта структура сшивается определенным образом (химически или ультрафиолетом), и дальше готовятся комплементарные зонды с остатками биотина, которые будут комплиментарны этой РНК. После того, как все сшили, подробили хроматин, к нему добавили биотинилированные зонды (на конце имеется модифицированный биотином азотистое основание). С биотином хорошо взаимодействует стрептавидин. За счет этого взаимодействия можно вытащить эти комплексы, удалить все кроме ДНК и определить нуклеотидную последовательность ДНК и смотреть, куда это все присоединится.
Как Linc узнают хроматин? Этот механизм напоминает узнавание ДНК сиквенс-специфичными белковыми факторами транскрипции.
• HOTAIR linc RNA предпочтительно занимают GA caйты (<500 нп) независимо от EZH2 (каталитическая субъединица для Н3K27me3)
• привлекается PcG комплекс, распространяющийся в обе стороны от сайта связывания linc
HOTAIR любит полипуриновые участки ДНК. Как она узнает ДНК: либо узнает триплексные структуры, либо узнает одну из нитей ДНК, это пока неясно. Единственное, когда первые данные об узнавании этой нкРНК были получены, сделали вывод, что этот механизм напоминает узнавание ДНК белковыми сиквенс-специфическими факторами, которые узнают определенные последовательности. Было еще выяснено, что для этого узнавания всякие комплексы белковые не нужны, а именно сама РНК узнает, а каталитические субъединицы не нужны. Когда узнавание нкРНК вместе с комплексом осуществилось, то сигнал распространяется в обе стороны от фокуса связывания.
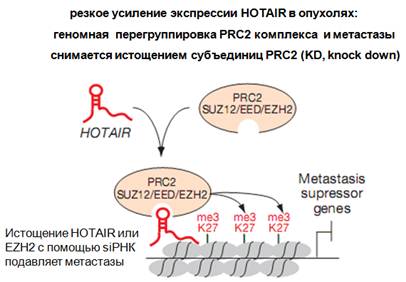
В опухоли, в частности молочной железы, HOTAIR экспрессируются в сотни, иногда даже тысячи раз сильнее, чем в нормальных неопухолевых в клетках. Причем оказалось, что ее экспрессия способствует тому, что меняется хроматиновый ландшафт клетки, то есть определенные гистоновые модификации. Оказалось, что привлекается комплекс PRC2, который осуществляет модификацию, причем характер их распределения меняется в присутствии избытка HOTAIR. Метилирование захватывает регуляторные участки и сами гены, которые являются супрессорами метастазирования. Активная экспрессия HOTAIR приводит к тому, что такие метки активны при метастазировании. Существуют способы детекции метастазирования: на определенную матрицу помещаются клетки и следят, как они делятся на матрице. Это все количественно оценивается. Считается, на основании полученных данных, что такая гиперэкспрессия HOTAIR осуществляет изменения эпигенома в опухолевой клетке, осуществляет его определенную модуляцию. При помощи siРНК показано, что если истощать HOTAIR или мРНК для каталитической субъединицы (делать нокдаун), то это будет подавлять метастазирование. Это имеет определенный клинический интерес. Осуществляется репрограммирование эпигеномного ландшафта.
экспрессия HOTAIR и метастазы
• Активация LINC в метастазирующих опухолях (в сотни-тысячи раз)
• Это приводит к репрограммированию профиля связывания PcG при раке груди
• Потеря HOTAIR подавляет инвазивность опухолевых клеток
• si RNA knockdown cубъединиц комплексов PcG cнижает метастазирование
• Функциональная взаимозависимость между HOTAIR и PRC2 комплексами
• Каковы механизмы, с помощью которых HOTAIR регулирует PRC2 комплексы? Не известно.
Перераспределение комплекса PRC.
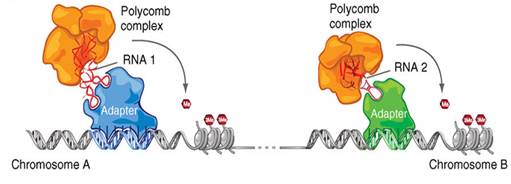
Может осуществляться перераспределение эпигеномных модификаций не только с помощью нкРНК и белковых комплексов, но и адаптерных белков, которые тоже узнают определенные участки ДНК, как транскрипционные факторы. Адаптерный комплекс имеет специфическое сродство к ДНК. Этот комплекс может участвовать в привлечении некой нкРНК и соответствующего PRC комплекса. Для того, чтобы разные РНК привлекались к разным участком ДНК, должны существовать разные адаптерные комплексы, которые взаимодействуют с разными нкРНК.
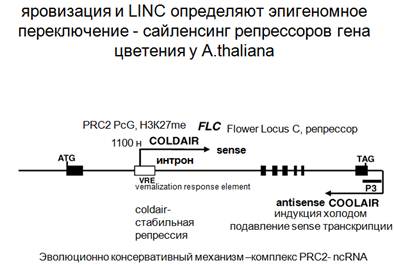
Удивительная консервативность: наличие комплексов PRC и наличие нкРНК для регуляции, которая имеет место быть не только у животных, но и у растений. Как было сказано ранее, siРНК которая определяет явление ко-супрессии и так далее, впервые была обнаружена у растений. Потом представления были перенесены на животные объекты. На A.thaliana исследовался вопрос регуляция цветения и яровизация. Яровизация - это выдерживание в холоде, которое в результате такого длительного воздействия приводит к изменению экспрессии, в данном случае к определенному (раннему) времени цветения А. thaliana. Здесь имеется ген, с экзон-интронным строением, типичное эукариотическое строение, когда огромные участки соответствует интронам. Этот ген дает некий транскрипционный фактор, который подавляет цветение. Этот фактор должен быть зарепрессирован и это делает холод. Наступает осень, зима, и начинает воздействие холод! Начинает образовываться антисмысловая РНК, Сoldair. По-видимому что-то происходит c хроматином, что обеспечивает изменение в хроматиновом состоянии в интроной части этого гена, откуда начинается транскрипция другой нкРНК в смысловом направлении (она не кодирует белок). Начинается эта транскрипция в интроне. Этот элемент генетически был выявлен как элемент, ответственный за яровизацию, за подавление экспрессия транскрипционного фактора. Оказалось, что это есть старт транскрипция Сoldair. Она привлекает гомологичный комплекс PRC2, который вносит метку, и благодаря этому осуществляется стабильная репрессия гена, который является репрессором цветения. И тогда цветение в определенный момент может начаться. Есть первая (нестабильная, начинающаяся) и вторая (стабильная) фаза яровизации.
Теперь интересный случай, когда нкРНК осуществляет активацию транскрипции. Есть репрессорные комплексы PRC, а есть активаторные, которые получили название tritorax по некой мутации. Если нарушаются соответствующие гены, которые действуют противоположно РRС, нарушается план строения тела дрозофилы. При нормальном развитии должен быть строгий регулируемый баланс между репрессорными и активаторными комплексами (между репрессорами этих генов и активаторами генов). Ранее было показано транс-действие репрессорного комплекса, а сейчас мы посмотрим цис-действие активаторного комплекса. Репрессорные (PcG) и активаторные (Trx – деметилирование Н3К27me3 и метилирование Н3К4me3) комплексы привлекаются как цис-, так и транс-действующими LINCs.
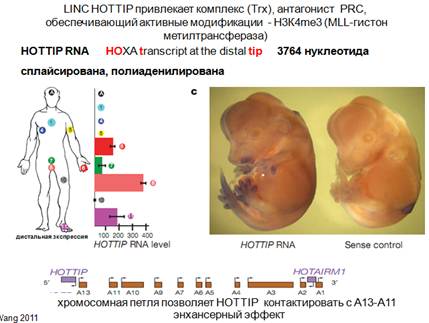
Смотрим рисунок. Низ - здесь представлен один из кластеров нох-генов под названием А. Кластер повернут на 180 градусов. Транскрипция в одном направлении. Как обычно, есть некая нкРНК (на общей схеме), которая транскрибируется в противоположном направлении от транскрипции генов этого кластера. Антисмысловая транскрипция в данном случае приводит к тому, что несколько генов цис-метилируются за счет этой транскрипции. Рассмотрим эпигеномное наследование на примере нкРНК, которая получила такое название hottip. На картинке транскрипция нох-генов у мышей, этот транскрипт у них транскрибируется на дистальных сторонах конечностей эмбриона. Проводилась детекция специфической hottip РНК с помощью гибридизации. И есть контроль на специфичность гибридизации (эмбрион справа). Показано, что hottip находится на дистальных частях конечностей. Если смотреть человека (фибробласты), то окажется, что этот hottip экспрессируется по разному. Столбики показывают интенсивность экспрессии и количество соответствующих транскриптов этой hot tip. Смотрим фиолетовый столбик: в дистальной части ступни там ее довольно много, в дистальной части руки ее тоже довольно много (кранный), а в туловище и проксимальный части практически ничего нет. Таким образом, имеется эпигеномное наследование, которое сохраняется и поддерживается этой hot tip.
Что еще интересного, каким образом распространяется достаточно далеко это активирующие влияние, как мы его можем регистрировать. Можно по эпигеномному ландшафту (для активации нужно чтобы была активная метка). Здесь представлено расположение соответствующих генов, которые активируется hottip. Изображена интенсивность метки, которую можно определенным образом оценивать (как в хроматине распределяется).
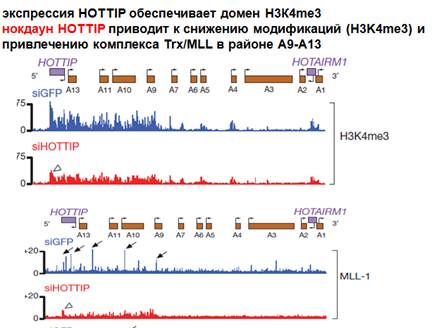
Метки очень много в районе, где транскрибируется сама hottip и ее достаточно много, где расположены гены, которые активны (активируюся). Теперь, если убрать hottip посредством siРНК, то получится сильное и достоверное снижение позитивных модификаций (красным обозначено). Видно очень мощное уничтожение этих модификаций самой hottip. Для того чтобы эта модификация возникала, как и в случае репрессорных комплексов, нужна не какая то одна субъединица, а весь комплекс. Одна из субъединиц этого комплекса, каталитическая, которая дает эту модификацию, связана с хроматином. Здесь имеется четкие пики в 5'-концах и межгенных промежутках, которые когда вы делаете нокдаун hottip, уничтожаются, значит это просто данные, которые показывают, что hottip заведует хроматином и хроматин эпигеномно наследуется при соматическом делении клетки. Как мы можем себе представлять эту активацию. Существуют методы: способы выяснения, какие геномные последовательности оказываются рядом друг с другом. Способ оценки конформации ДНК в геноме. 3С (chromosome conformation capture). Метод детекции определенной хромосомной конформации.
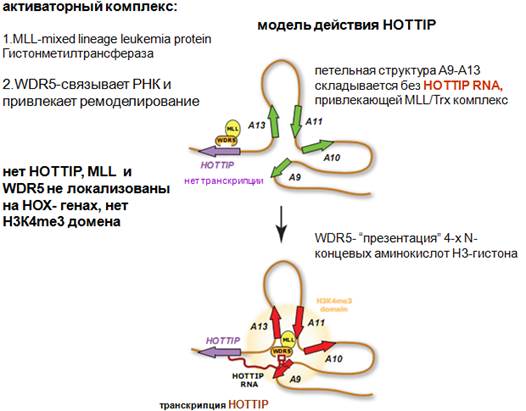 Когда стали исследовать хромосомную конформацию hottip и активирующихся генов, оказалось, что активирующиеся гены уложены в определенную петельную структуру, благодаря чему эти гены и hottip сближены в неком компартменте. Петельная структура складывается без hottip. Ждет, когда начнется транскрипция hottip. И когда она начинается, она привлекает активаторные комплексы, которые содержат несколько субъединиц. Назовем только две: МLL-mixed lineage leukemia protein - Гистонметилтрансфераза и WDR5-cвязывает РНК и привлекает ремоделирование. Содержит повторы АК, такие же были у белка, который вносил репрессорную модификацию у PCR2 комплекса. В данном случае, эти повторы связывают РНК. Если MLL не образуется, или поврежден ген, то будет лейкемия.
Когда стали исследовать хромосомную конформацию hottip и активирующихся генов, оказалось, что активирующиеся гены уложены в определенную петельную структуру, благодаря чему эти гены и hottip сближены в неком компартменте. Петельная структура складывается без hottip. Ждет, когда начнется транскрипция hottip. И когда она начинается, она привлекает активаторные комплексы, которые содержат несколько субъединиц. Назовем только две: МLL-mixed lineage leukemia protein - Гистонметилтрансфераза и WDR5-cвязывает РНК и привлекает ремоделирование. Содержит повторы АК, такие же были у белка, который вносил репрессорную модификацию у PCR2 комплекса. В данном случае, эти повторы связывают РНК. Если MLL не образуется, или поврежден ген, то будет лейкемия.
Что здесь получается. Когда белок WDR5 связывается с РНК, в составе комплекса осуществляется изменение конформации гистонов в составе нуклеосом. Этот белок осуществляет презентацию – подставляет для другой субъединицы гистонметилтрансферазы N-конец гистона, для того, чтобы она его метилировала. Подставляется несколько АК с N-конца. Структура представлена т.о., чтобы hottip привлекла еще связанный с WDR5 белок, который осуществляет модификацию хроматина, которая была показывана на хроматиновых ландшафтах на предыдущей картинке. Видим, что хроматиновые ландшафты достаточно протяженные, почему-то они существуют локусами. Считается, что благодаря протяженности, осуществляется стабильность хромосомных ландшафтов. Эти структуры между собой взаимодействуют и обеспечивают стабильность.
Что еще интересного, относительно LINC RNAs. Роль: Клеточная пролиферация и p53, Дифференцировка, Плюрипотентность (“cтволовость”). Будем смотреть за linc, которая зависит от р53 – TF, который узнает определенные последовательности ДНК и активирует экспрессию. Но наряду с этим он может осуществлять репрессорные функции (косвенно, активируя репрессорный белок). Также его репрессорная функция может осуществляться в том, что он, как активатор, индуцирует транскрипцию linc. И вообще промоторы заметного количества нкРНК содержат сайты р53. Промоторы нкРНК содежат сайты для ТF, определяющие стволовость.
Транс-действующие linc RNA, зависимые от р53, свойства:
• Активаторные и репрессорные функция р53 как транскрипционного фактора хорошо известны и активно исследуются
• Репрессорная функция р53 может осуществляться благодаря индукции транскрипции lincРНК
• Промоторы нкРНК содержат сайты для p53
Мы не знаем, что делает нкРНК, но видим, что она активируется такими-то факторами. Посмотрим на Linc р21, она соседствует с геном, кодирующим р21.
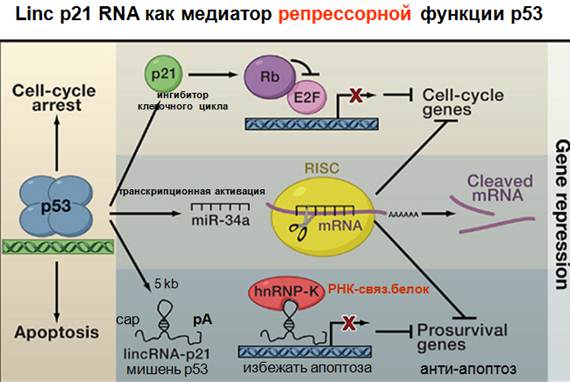
Белок р21 известен тем, что является ингибиторам cdk, клеточного цикла, останавливает его, является антионкогеном. Представлен как фактор, соединяющийся с ДНК. Р53 его регулирует. Здесь показана транскрипционная активация. Также р53 способствует транскрипции нкРНК 5kb, которая имеет кэп, полиА, т.е. нормальная РНК, но некодирующая. И эта нкРНК в составе комплекса способствует тому, чтобы клетки скорее входили в апоптоз. Ген, подавляемый комплексом, работает как стимулятор выхода из апоптоза и стимулятор выживания клетки и выхода ее из деления. Этот комплекс р53 это подавляет, и в этом заключается антионкогенная функция белка р53. Теперь по поводу открытой недавно linc, которая нужна для дифференцировки клеток. В данном случае, для дифференцировки клеток кожи. Кератиноцитов. Во главе исседований – китаец, который раньше был дерматологом.

Дифференцировка клеток кожи. Если дифференцировки не происходит, то появляются разнообразные болезни. Дифференцировку клеток кожи надо исследовать, надо понимать, что это такое. Была обнаружена некая нкРНК, достаточно длинная, около 4 kb. Цитоплазматическая. До этого мы с вами разбирали ядерные, которые действовали на уровне хроматина. Длинные цитоплазматические РНК тоже оказывают интересные действия. Играет ключевую роль в терминальной дифференцировке. Что показано на этой диаграмме. Здесь отложена условно экспрессия почти трехсот генов, вернее определенных транскриптов, которые образуются до дифференцировки. Синим показано, что их мало, красным показано, что их много. (В определенной логарифмической шкале). Если посчитать эти полосочки, то насчитаем 258 штук. Дальше, когда начинается дифференцировка, количество транскриптов, которых очень мало (синие), резко увеличивается, а потом резко падает. Что происходит с этой нкРНК. Ее практически нет до дифференцировки, потом добавляем кальций, предшественник кератиноцитов, и оказывается, что появляется нкРНК ТINCR, а потом ее становится значительно больше. Все эти данные получены по новой технологии секвенирования РНК: вы кидаете в общий биоинформатический пул, а потом из него возникают данные. Как ТINCR может действовать, и как она может определять дифференцировку? Оказалось, что составе этой ТINCR имеется нуклеотидные последовательности из 25 нуклеотидов, которые могут взаимодействовать с комплементарной последовательностью в информационных белок-кодирующих РНК и тем самым стабилизировать эти РНК. Как стабилизировать, это остается непонятно. Высказывалось предположение, что нкРНК могут отсасывать на себя как губки микроРНК, которые подавляют экспрессию мРНК тем или иным способом. Здесь показано, что ТINCR не содержит последовательностей комплементарных микроРНК, то есть она не является comparative endogenous РНК. Губки конкурируют за микроРНК, для того чтобы обеспечить трансляцию соответствующих мРНК.
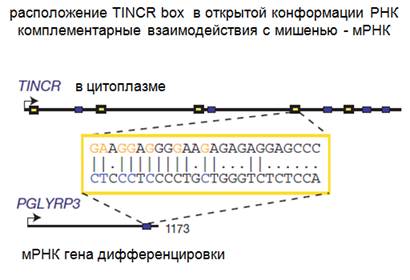
Здесь показано расположение ТINCR. Имеется некая мишень, это мРНК, которая нужна для дифференцировки. Она имеет участок комплиментарный боксу и счет этого осуществляется ее стабилизация. Если соответствующий бокс будет удален из РНК, ее присутствие падает во много раз. Этот эксперимент показывает, каким интересным способом действует нкРНК на уровне трансляции, механизм стабилизации спрашивайте у лекторов, которые читают механизм трансляции. Должно быть исследовано. А теперь про эмбриональные стволовые клетки и влияние нкРНК.
LINC and ESCs (embryonicl stem cells)
• Knockdown pяда Linc приводит к выходу из плюрипотентного состояния (тот же эффект, что и КD мРНК рег-ых белков)
• Linc, не требуемые для поддержания плюрипотентности, являются репрессорами клеточной дифференцировки
• Linc RNAs регулируются транскрипционными факторами плюрипотентности
• Взаимодействуют со специфичными регуляторными хроматиновыми комплексами
• В основном trans-действие: нокдаун оказывает влияние не только на близлежащие гены
Чтобы доказать, что они очень важны для стволовых клеток, делается нокдаун siРНК, и клетка лишается плюрипотентного состояния. Тоже самое, если вы сделаете нокдаун мРНК, кодирующей белок плюрипотентности. Эти linc регулируются транскрипционными факторами плюрипотентности. Взаимодействуют со специфичными регуляторными хроматиновыми комплексами. Для этих плюрипотентных linc характерно транс-действие. Если вы делаете нокдаун этой нкРНК, то эффект вы получаете на множество генов, не в одном месте, где она транскрибируется. Т.е. имеется транс-действие. Механизм не исследован.
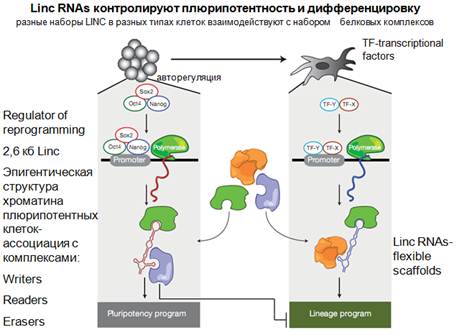
Здесь представлена сумма современных знаний. С помощью факторов плюрипотентности получается индуцированные плюрипотентные клетки. Они осуществляют репрограммирование искусственно, запускают транскрипция нкРНК, которые имеют такой скелет, который взаимодействует с определенными белковыми факторами и определяет программу плюрипотентности. Здесь интересно, что эти комплексы еще действуют на промоторы генов, которые определяют эти факторы. Это стимулирующее действие, за счет этого осуществляется авторегуляция стволовых клеток, образование этих TF. Справа показана дифференцированная клетка. И там образуются другие нкРНК, с другим гибким скелетом, на котором формируются некие другие комплексы, которые уже запускают не программу плюрипотентности, а какую-то другую программу, скажем дифференцировки в определенные клеточные линии. И для этой, и для этой программы может использоваться некий общий пул комплексов белковых, комбинация которых может приводить к совсем разным эффектам.
Геномный родительский импринтинг.
· Подавляется экспрессия генов на одной из гомологичных хромосом (материнский или отцовский импринтинг)
· «Гаплоидная», или моноаллельная экспрессия гена определяется наследованием гена от отца или от матери
· Импринтированные гены кодируют белки, регулирующие эмбриональное и неонатальное развитие
· Импринтинг определяется метилированием ДНК
· Динамика метилирования и деметилирования в развитии
Особая ролт отводится linc, которые имеют отношение к метилированию ДНК. Механизмы метилирования с участием нкРНК у животных изучены плохо. Геномный родительский импринтинг заключается в том, что в некоторых случаях наблюдается экспрессия генов только с одной из гомологичных хромосом. В одном случае только с материнской хромосомы, в другом только с отцовской. По видимому, у плацентарных и млекопитающих в эволюции появился этот геномный импринтинг. У человека имеется сотни генов, которые так себя ведут. Вначале это было показано, когда пересаживали пронуклеусы. После оплодотворения отцовское и материнское ядра еще не слились, и тогда можно их вытащить, пересадить и посмотреть, что происходит, если мы будем сливать 2 материнских ядра или 2 отцовских. Что же получается? Ничего хорошего не получается. Если сливаются 2 материнских, то получается зародыш (не очень нормальный, но получается) без оболочек. Если взять два мужских, зародыша практически нет, но оболочки есть. Для некоторых генов должно быть удивительное соответствие между активностью аллельных генов, должна соблюдаться некая доза этой активности, она не должна быть в некоторых случаях очень высокой, а должна держаться на определенном уровне. Т.е. для сотни генов имеется «гаплоидная», или моноаллельная экспрессия генов, такие гены называют импринтированными. Это гены, кодирующие белки, которые регулируют эмбриональное развитие (в основном и после рождения самый первый этап развития). Это достаточно важный процесс, потому что при его нарушении происходят различные аномалии. Импринтинг определяется метилированием определенного участка ДНК, соседствующего с геном, который подвергается репрессии. Метилирование ДНК это динамичный процесс который меняется в развитии. Рассмотрим в общем, что это такое.
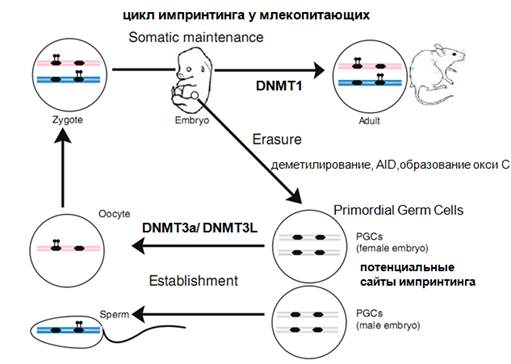
Вот есть две гомологичные хромосомы: отцовская и материнская. Шарики-леденцы показывают, что произошло метилирование, а в гомологичной хромосоме этого нет. Это ряд относительно небольшого числа генов. Дальше начинается развитие. Происходит деметилирование, которое не захватывает метилирование импринтированных генов. Дальше возникает новое метилирование в развитии, не импринтированных генов, а разных генов. А импринтированные гены не подвергаются деметилированию, которое происходит в развитии. Они устойчивы. У животных на самых ранних стадиях развития обособляются клетки, которые дадут ткань зародышевого пути. Здесь будет осуществляться деметилирование импринтированных генов. Если в соматических тканях оно сохранялось, то здесь оно будет стираться. Т.о. в предшественниках герминальных клеток имееются потенциальный сайты метелирования (у предшественников ооцитов и спермы). И дальше с участием ДНК-метилтрансфераз осуществляется метелирование, которое будет различаться по своему рисунку. И это все будет сохраняться при оплодотворении и в дальнейшем развитии. Прежде всего, было выяснено, что импринтированные гены стремятся находиться в кластерах, одиночки есть, но их мало.
Кластеризация импринтированных генов
• 3-10 генов вместе с геном noncoding RNA (lncRNA) в кластере (домен)
• Lnc RNA образуется на одной из родительских хромосом - сайленсинг хромосомного домена
• Цис-действие noncoding RNA
• IC (imprinting control) element/region (ICE или ICR)
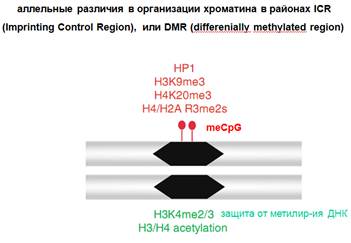 Например, несколько генов находятся в одном кластере, и в этом же кластере находится ген, транскрипция которого дает длинную нкРНК. Считается, что этот кластер из белок-кодирующих генов и нкРНК образуют некий домен, который может быть активным или неактивным для белок-кодирующих генов, которые находятся в этом кластере. И дальше оказывается, что длинная нкРНК образуется только на одной из родительских хромосом (для импринтированных генов) и осуществляет сайленсинг этого хромосомного домена. И это есть цис-действие, т.е. компактизация распространяется только на эти кластеры. Дальше в этом кластере выявляются контролирующие элементы, Imprinting Control Region. Это те точки, которые определяют метилирование ДНК. ICR могут находиться в альтернативных состояниях: могут быть метилированными или неметилированными, поэтому их называют дифференциально метилированными участками (в зависимости от характера импринтинга). Они показаны на картинке. Это два гомологичных участка. Они унаследованы от разных родителей. Один из них метилирован. Показаны сопутствующие репрессорные метки хроматина (хроматиновый белок НР1, и т д). А в участке на гомологичной хромосоме – активные модификации. В частности, H3K4me2/3 защищает от метилирования ДНК. Только если деметилаза ее уберет, то осуществиться метелирование ДНК. Это очень важный момент регуляции. Ведь впервые на растениях было показано, что состояние белков хроматина может определять метилирование ДНК.
Например, несколько генов находятся в одном кластере, и в этом же кластере находится ген, транскрипция которого дает длинную нкРНК. Считается, что этот кластер из белок-кодирующих генов и нкРНК образуют некий домен, который может быть активным или неактивным для белок-кодирующих генов, которые находятся в этом кластере. И дальше оказывается, что длинная нкРНК образуется только на одной из родительских хромосом (для импринтированных генов) и осуществляет сайленсинг этого хромосомного домена. И это есть цис-действие, т.е. компактизация распространяется только на эти кластеры. Дальше в этом кластере выявляются контролирующие элементы, Imprinting Control Region. Это те точки, которые определяют метилирование ДНК. ICR могут находиться в альтернативных состояниях: могут быть метилированными или неметилированными, поэтому их называют дифференциально метилированными участками (в зависимости от характера импринтинга). Они показаны на картинке. Это два гомологичных участка. Они унаследованы от разных родителей. Один из них метилирован. Показаны сопутствующие репрессорные метки хроматина (хроматиновый белок НР1, и т д). А в участке на гомологичной хромосоме – активные модификации. В частности, H3K4me2/3 защищает от метилирования ДНК. Только если деметилаза ее уберет, то осуществиться метелирование ДНК. Это очень важный момент регуляции. Ведь впервые на растениях было показано, что состояние белков хроматина может определять метилирование ДНК.

Теперь об импринтинге двух структурных кластеров генов. Нас интересует ген, кодирующий фактор роста, зависящий от инсулина. Этот ген очень важен для раннего развития эмбриона. Считается, что будет очень плохо, если этот ген унаследуется в двух копиях – и от отца, и от матери в активном состоянии. Этот приведет к тому, что рост эмбриона будет очень интенсивный, плацента и репродуктивные органы матери будут изнашиваться. Это объясняет, для чего нужен импринтинг, т.е. подавление фактора роста в отцовском аллеле. В отцовском аллеле в контролирующем районе не происходит метилирования. Хроматин защищен от метилирования меткой. И отсюда начинается транскрипция антисмысловой межгенной нкРНК. Она каким-то образом осуществляет сайленсинг не только этого гена, но и генов, находящихся по другую сторону старта транскрипции. Если сделать некие искусственные трансгенные конструкции и делать так, чтобы транскрипция в месте нкРНК оборвалась, то импринтинга не будет. Как осуществляется подавление, неясно. Представляется, что есть некий домен за счет того, что имеется определенная укладка ДНК и привлечение факторов, регулирующих транскрипцию и состояние хроматина, осуществляется за счет нкРНК. Надо сказать, что подавление крайне специфично: во всех тканях подавлен один ген, а только в плаценте подавлен другой ген. Аналогичная история для гена, кодирующего кальциевый канал (внизу этой картинки). В интроне гена находится район, контролирующий метилирование нкРНК. Длинная нкРНК начинает транскрибироваться с интрона и подавляет экспрессию многих генов, находящихся в кластере. Можно думать, что какие-то транскрипты дают дцРНК, а далее siРНК…Но это не так! Показано, что дайсер здесь никак не участвует. Т.е. сайленсинг осуществляется без siРНК, просто linc организуют каким-то образом домен, который организует двунаправленный сайленсинг.
Регуляция: ICE (IC, ICR) imprint control element.
• Метилирование ICE в гаметах запрещает образование lnc RNA и обеспечивает экспрессию генов кластера
• Неметилированный ICE позволяет экспрессироваться lnc RNA, вызывая цис- сайленсинг генов кластера, кодирующих белки
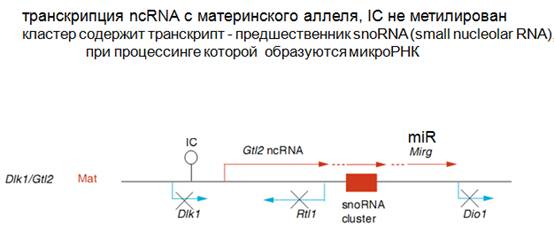
Рассмотрим другой случай, когда транскрипт с материнского аллеля неметилирован. (Выше было о том, что с отцовского аллеля неметилирован). Образуется некая нкРНК, которая осуществляет подавление экспрессии определенных генов. Опять тут транскрипция генов двунаправленная. Что еще интересно. С одной стороны, длинная нкРНК нужна для создания структуры этого домена. С другой стороны, оказывается, что в результате процессинга этой нкРНК образуются малые ядерные РНК, которые необходимы для модификации рибосомных РНК, РНК для сплайсинга, т.е. это очень важные РНК для всех клеток. И среди них есть miR. На рисунке показано, как один и тот же транскрипт может иметь несколько разных функций.
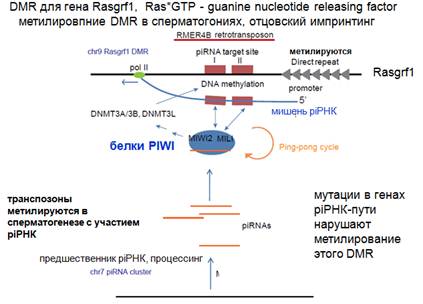
Здесь показан довольно необычный случай импринтинга. Ген достаточно важный, определяет связывание ГТФ с Ras (протоонкоген, участвующий в сигнальном пути). Когда Ras связан с ГДФ, то продукт гена способствует освобождению ГДФ из Ras, затем Ras снова может воспринять ГДФ и дальше участвовать в сигнальном пути. Имеется ген и имеется район, который контролирует импринтинг. Ген будет работать в том случае, если участок (район) будет метилирован, и если с него не будет образовываться в заметном количестве нкРНК. Этот участок дифференциально метилированный: в хромосоме, идущей от матери, он не метилирован, в отцовской метилирован. Имеется район, состоящий из прямых повторов, которые метилируются, дальше район, который содержит последовательность транспозона, экспрессия которого подавляется piРНК (транспозон метилируется) и осуществляется транскрипция этого участка. А промотором для транскрипции служит участок с прямыми повторами. Что происходит в случае отцовского происхождения этого участка. В хромосоме 9 имеется этот ген, а в другой хромосоме имеется кластер piРНК, с которых они образуются и определенным образом процессируются, эти piРНК с участием белков PIWI могут осуществлять процессинг новообразованных РНК, и здесь может осуществляться что-то вроде пинг-понг цикла, приводящего к амплификации piРНК. Дальше, что происходит. PiРНК, которая накапливается в результате этих процессов, узнает новообразованный транскрипт, а может быть ДНК, и привлекает ДНК-метилтрансферазу, которая все метилирует. Тогда эта нкРНК не образуется (только в зачаточном состоянии). И ее недостаточно, чтобы образовывать домен для сайленсинга гена. Т.е. этот ген в активном виде наследуется только с отцовской хромосомы. Если вы получаете мутацию по piРНК сайленсингу, то нарушается все метилирование. Если получить делецию по промотору и нарушить транскрипцию, то это тоже нарушает метилирование. Т.е. мы приходим к представлению о том, что для метилирования оказывается нужна транскрипции, это похоже на случай у растений, когда полимераза V осуществляла транскрипцию, и это было нужно для метилирования ДНК.
|
|
|
|
Дата добавления: 2014-01-03; Просмотров: 1248; Нарушение авторских прав?; Мы поможем в написании вашей работы!