КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Лекция № 4. Тема:Биоэлектрогенез. Электрические и магнитные свойства тканей и окружающей среды
|
|
|
|
Тема: Биоэлектрогенез. Электрические и магнитные свойства тканей и окружающей среды
План лекции:
1. Мембранные потенциалы и их ионная природа. Потенциал покоя. Механизм генерации и распространения потенциала действия
2. Понятие диполя. Диполь в электрическом поле
3. Токовый диполь. Представление об эквивалентном электрическом генераторе органов и тканей
4. Генез электрокардиограммы в рамках модели дипольного эквивалентного электрического генератора
1. Одна из важнейших функций биологической мембраны - генерация и передача биопотенциалов. Это явление лежит в основе возбудимости клеток, регуляции внутриклеточных процессов, работы нервной системы, регуляции мышечного сокращения, рецепции. В медицине на исследование электрических полей, созданных биопотенциалами органов и тканей, основаны диагностические методы: электрокардиография, электроэнцефалография, электромиография и другие. Практикуется и лечебное воздействие на ткани и органы внешними электрическими импульсами при электростимуляции.
В процессе жизнедеятельности в клетках и тканях могут возникать разности электрических потенциалов:
1) окислительно-восстановительные потенциалы - вследствие переноса электронов от одних молекул к другим;
2) мембранные - вследствие градиента концентрации ионов и переноса ионов через мембрану.
Биопотенциалы, регистрируемые в организме,- это в основном мембранные потенциалы.
Мембранным потенциалом называется разность потенциалов между внутренней (цитоплазматической) и наружной поверхностями мембраны.
Мембранная разность потенциалов рассчитывается по формуле Нернста:
Δ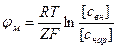 , (1)
, (1)
где свх и снар — молярные концентрации ионов по обе стороны мембраны, R — универсальная газовая постоянная, T — термодинамическая температура, при которой происходит диффузия, F - постоянная Фарадея, Z — заряд иона. Эту разность потенциалов называют равновесным мембранным потенциалом. Мембранная теория происхождения биопотенциалов была выдвинута в 1902 г. Б. Бернштейном. Действительно, в живой клетке концентрация ионов калия значительно больше, чем в межклеточной жидкости, и крупные органические молекулы практически проникают через мембрану. Важным доводом в пользу представлений Бернштейна послужил тот факт, что рассчитанная по формуле Нернста разность потенциалов между наружной и внутренней сторонами мембраны мышечного волокна оказалась близкой к измеренной в опытах с помощью внутриклеточного микро электрода.
Однако «калиевая теория» мембранного потенциала оказалась несовершенной, не способной объяснить наблюдаемые впоследствии факты отклонения истинных значений потенциалов на мембранах живых клеток от теоретически рассчитанных. Оказалось, что равновесный мембранный потенциал характерен лишь для мертвых клеток, либо клеток с ослабленным метаболизмом.
В настоящее время общепризнанной теорией, объясняющее возникновение и поддержание потенциала на клеточной мембране в состоянии физиологического покоя, является теория А. Ходжкина. Она была развита и экспериментально обоснована им в 50-х гг. XX в. Сущность ее заключается в том, что потенциал, существующий на мембранах невозбужденных клеток (потенциал покоя) обусловлен полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и окружающей средой. Это распределение поддерживается механизмами активного переноса, локализованными в самой мембране.
Потенциал покоя – стационарная разность электрических потенциалов, регистрируемая между внутренней и наружной поверхностями мембраны в невозбужденном состоянии.
Учтя влияние на величину потенциала покоя и ионов натрия и хлора, в 1949 г. Гольдман, Ходжкин и Катц вывели для величины потенциала покоя уравнение (оно называется уравнением Гольдмана –Ходжкина - Катца):
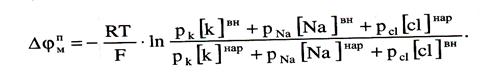 (2)
(2)
Оно было получено из того предположения, что в невозбужденном состоянии через поверхность мембраны проходят потоки ионов натрия и хлора, алгебраическая сумма которых должна быть нулю:
jK + jNa + jCl = 0. (j – плотность потока вещества, [моль/м2.с].
Очевидно, если в этом уравнении вновь пренебречь проницаемостью мембраны для ионов натрия и хлора по сравнению с проницаемостью для иона калия, оно автоматически станет уравнением Нернста.
Для живой клетки характерно не равновесное распределение веществ, но существование потоков ионов в обе стороны через мембрану. Такое состояние, при котором число различных ионов, проходящих в единицу времени через мембрану внутрь клетки, равно числу выходящих из клетки ионов, называют стационарным. Ясно, что в стационарном состоянии суммарная плотность потока ионов через мембрану равна нулю: j = 0.
Различные концентрации ионов внутри и вне клетки созданы ионными насосами — системами активного транспорта. Можно сказать, что потенциал покоя обязан активному переносу.
Известно, что проницаемость мембраны для ионов калия самая высокая. В состоянии покоя соотношение коэффициентов проницаемости для разных ионов равно:
PK = PNa = PCl = 1: 0.04: 0.45, (3)
где P = DK/l [м/с], К – коэффициент распределения, который показывает соотношение концентрации вещества вне мембраны и внутри ее, D – коэффициент диффузии (D = URT), U – подвижность частиц).
Вследствие этого диффузия калия и хлора идет в обе стороны. Натрий идет через мембрану за счет простой диффузии лишь в одну сторону — снаружи вовнутрь. Однако Nа+-К+-АТФаза интенсивно выводит ионы натрия из клетки, а калия — в клетку. Потенциал покоя, рассчитанный по формуле Гольдмана—Ходжкина- Катца, составляет 80 мВ со знаком минус со стороны внутриклеточного пространства.
Ходжкин и Хаксли сформулировали гипотезу о том, что при возбуждении меняется характер проницаемости мембраны для разного типа ионов: из проницаемой главным образом для калия она превращается в проницаемую для натрия и, следовательно, поток ионов натрия внутрь клетки увеличивается. Разность потенциалов между поверхностями мембраны поднимается до нуля, а затем становится положительной — наступает деполяризация мембраны. Далее процесс развивается в обратном направлении (процесс реполяризации): поток ионов натрия внутрь клетки постепенно уменьшается и возвращается к «невозбужденному» значению; разность потенциалов также возвращается к значению Δφм. Такой импульс (рис. 1) называют потенциалом действия – э лектрический импульс, обусловленный изменением ионной проницаемости мембраны и связанный с распространением по нервам и мышцам волны возбуждения.
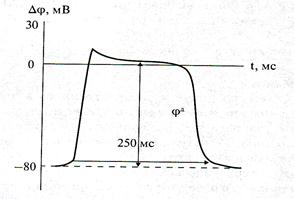
Рис.1
Возбужденный импульс вызывает лишь на короткое время смещение мембранного потенциала, которое быстро пропадает и восстанавливается потенциал покоя, в том случае, когда возбуждающий импульс отрицательный – приводит к изменению мембранного потенциала еще лальше в отрицательную сторону, то есть возбуждающий импульс гиперполяризующий. Также не формируется потенциал действия, когда возбуждающий импульс положительный (деполяризующий), но его амплитуда меньше порогового значения Vпор. Однако, если амплитуда положительного деполяризующего импульса окажется больше значения Vпор, то φм становится больше φмпор и в мембране развивается процесс, в результате которого происходит резкое повышение мембранного потенциала и мембранный потенциал φм даже меняет свой знак — становится положительным (φвн > φнар).
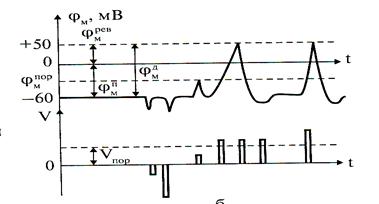
Рис.2
Достигнув некоторого положительного значения φмрев — потенциала реверсии, мембранный потенциал возвращается к значению потенциала покоя φмп, около этого значения наблюдаются затухающие колебания. В нервных волокнах и скелетных мышцах длительность потенциала действия около 1 мс (а в сердечной мышце около 300 мс). После снятия возбуждения еще в течение 1—3 мс в мембране наблюдаются некоторые остаточные явления, во время которых мембрана рефрактерна (невозбудима) (см. Рис.2).
Новый деполяризующий потенциал V > Vпор может вызвать образование нового потенциала действия только после полного возвращения мембраны в состояние покоя. Причем амплитуда потенциала действия
φм д = ׀φмп ׀+ φмрев (4)
не зависит от амплитуды деполяризующего потенциала, если только V > Vпор
Если в покое мембрана поляризована, - потенциал цитоплазмы отрицателен по отношению к внеклеточной среде, то при возбуждении происходит деполяризация мембраны — потенциал внутри клетки положителен и после снятия возбуждения происходит реполяризация (восстановление поляризации) мембраны. Характерные свойства потенциала действия:
1) наличие порогового значения деполяризующего потенциала;
2) закон «все или ничего», то есть, если деполяризующий потенциал больше порогового, развивается потенциал действия, амплитуда которого не зависит от амплитуды возбуждающего импульса, и нет потенциала действия, если амплитуда деполяризующего потенциала меньше пороговой;
3) есть период рефрактерности (невозбудимости мембраны) во время развития потенциала действия и остаточных явлений после снятия возбуждения;
4) в момент возбуждения резко уменьшается сопротивление мембраны (у аксона кальмара от 0,1 Ом • м2 в покое, до 0,0025 Ом • м2 при возбуждении).
Если обратиться к данным для значений равновесных нернстовских потенциалов, созданных различными ионами, естественно предположить, что положительный потенциал реверсии имеет натриевую природу, поскольку именно диффузия натрия создает положительную разность потенциалов между внутренней и наружной поверхностями мембраны.
Можно менять амплитуду импульса потенциала действия, изменяя концентрацию натрия в наружной среде. При уменьшении наружной концентрации натрия амплитуда потенциала действия уменьшается, так как меняется потенциал реверсии. Если из окружающей клетку среды полностью удалить натрий, потенциал действия вообще не возникает.
Установлено, что формирование потенциала действия вызывается ионными потоками через мембрану: сначала ионов натрия клетки, а затем — ионов калия из клетки в наружный раствор связано с изменением проницаемости мембраны для ионов к натрия.
Распространение нервного импульса вдоль возбудимого волокна происходит следующим образом.
Если в каком-нибудь участке возбудимой мембраны сформировался потенциал действия, мембрана деполяризована, возбуждение распространяется на другие участки мембраны. Рассмотрим распространение возбуждения на примере передачи нервного импульса по аксону (рис. 3).
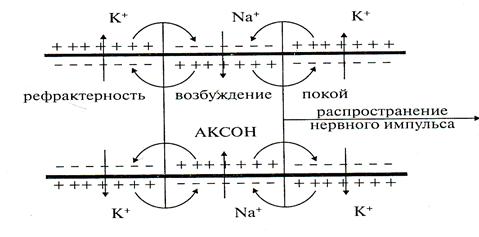
Рис.3
И в аксоплазме, и в окружающем растворе возникают локальные токи: между участками поверхности мембраны с большим потенциалом (положительно заряженными) и участками с меньшим потенциалом (отрицательно заряженными).
Локальные токи образуются и внутри аксона, и на наружной его поверхности. Локальные электрические токи приводят к повышению потенциала внутренней поверхности невозбужденного участка мембраны φвн и к понижению φнар — наружного потенциала невозбужденного участка мембраны, оказавшегося по соседству с возбужденной зоной. Таким образом, отрицательный потенциал покоя φмп уменьшается по абсолютной величине, то есть повышается. В областях, близких к возбужденному участку, φм повышается выше порогового значения. Под действием изменения мембранного потенциала открываются натриевые каналы и дальнейшее повышение происходит уже за счет потока ионов натрия через мембрану.
Происходит деполяризация мембраны, развивается потенциал действия. Затем возбуждение передается дальше на покоящиеся участки мембраны.
Разность потенциалов между отдельными участками клетки или волокна может быть зарегистрирована с применением микроэлектродной техники. При функционировании тканей и органов, как и отдельных клеток, сопровождающемся электрической активностью, в организме обнаруживается электрическое поле.
2. В период деполяризации и реполяризации наружная поверхность мембраны как бы состоит из двух полюсов – положительного и отрицательного.
Электрическая система, образованная двумя равными по величине, но противоположными по знаку зарядами, расположенными на малом расстоянии друг от друга, называется диполем.
Основной характеристикой диполя (Рис.4) является его электрический момент (дипольный момент) – вектор, равный произведению заряда q на плечо диполя l, направленный от отрицательного заряда к положительному:
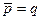
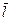 (5)
(5)

Рис. 4
Поместим диполь в однородное электрическое поле напряженностью 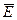 .
.
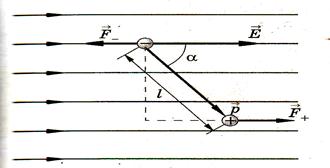
Рис.5
На каждый из зарядов диполя действуют силы 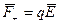 и
и  , эти силы равны по модулю, противоположно направлены и создают момент пары сил: M = q E l sin ά = p E sin ά, или в векторной форме
, эти силы равны по модулю, противоположно направлены и создают момент пары сил: M = q E l sin ά = p E sin ά, или в векторной форме 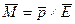 .
.
Таким образом, на диполь в однородном электрическом поле действует момент силы, зависящий от электрического момента и ориентации диполя, а также напряженности поля.
Поместим диполь в неоднородное электрическое поле. Предположим, что диполь расположен вдоль силовой линии (рис.6).
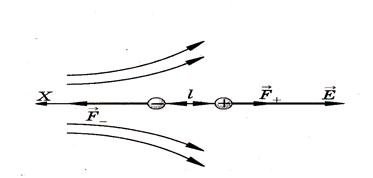
Рис.6
На диполь действует сила, зависящая от его электрического момента и степени неоднородности поля dE/dx: F = p dE/dx.
3. До сих пор рассматривался диполь, помещенный в электрическое поле, однако сам диполь также является источником поля.
Пусть диполь, создающий электрическое поле, находится в центре равностороннего треугольника АВС (рис. 7а, б). Тогда можно получить, что напряжения на сторонах этого треугольника относятся как проекции вектора 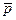 на его стороны:
на его стороны:
UАВ: UВС: UСА = pАВ: pВС: pСА (6)
Рис.7а
Исследуя изменения разности потенциалов на поверхности человеческого тела, можно судить о проекциях дипольного момента сердца, следовательно, о биопотенциалах сердца.
В вакууме или в идеальном изоляторе электрический диполь может сохраняться сколь угодно долго. Однако в реальной ситуации (электропроводящая среда) под действием электрической: поля диполя возникает движение свободных зарядов и диполь либо экранируется, либо нейтрализуется.
Можно к диполю подключить источник напряжения, иными словами, клеммы источника напряжения представить как диполь. В этом случае, несмотря на наличие тока в проводящей среде, диполь будет сохраняться (рис. 8). Резистор R1 является эквивалентом сопротивления проводящей среды, ε — ЭДС источника, r — его внутреннее сопротивление (рис. 9).
Рис. 8
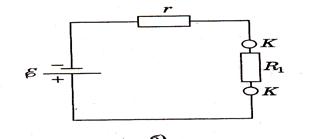
Рис.9
На основании закона Ома для полной цепи I = ε / R + r
если г >> R1, то I = ε / г.
Можно заключить, что в этом случае сила тока во внешней цепи будет оставаться почти постоянной, она почти не зависит от свойств среды (при условии г >> R1. Такая двухполюсная система, состоящая из истока и стока тока, называется дипольным электрическим генератором или токовым диполем.
Между дипольным электрическим генератором и электрическим диполем имеется большая аналогия, которая основывается на общей аналогии электрического поля в проводящей среде и электростатического поля.
Суть аналогии электрического поля в проводящей среде и электростатического поля сводится к следующему:
— линии тока (электрическое поле в проводящей среде) совпадают с линиями напряженности электростатического поля при одинаковой форме электродов;
— в том и другом случаях многие формулы имеют тождественный вид.
Воспользуемся этой аналогией и получим выражение для токового диполя. Аналогично электрическому моменту диполя введем дипольный момент дипольного электрического генератора:
Pт = I l,
где l — расстояние между точками истока и стока тока.
В соответствии с изложенным можно ввести и понятие мультипольного электрического генератора.
По существу, электрический мультипольный генератор — это некоторая пространственная совокупность электрических токов (совокупность истоков и стоков различных токов).
4. Живые ткани являются источником электрических потенциалов (биопотенциалов).
Регистрация биопотенциалов тканей и органов с диагностической (исследовательской) целью получила название электрографии. Такой общий термин употребляется сравнительно редко, более распространены конкретные названия соответствующих диагностических методов: электрокардиография (ЭКГ) — регистрация биопотенциалов, возникающих в сердечной мышце при ее возбуждении, электромиография — метод регистрации биоэлектрической активности мышц, электроэнцефалография (ЭЭГ) — метод регистрации биоэлектрической активности головного мозга и др.
В большинстве случаев биопотенциалы снимаются электродами не непосредственно с органа (сердце, головной мозг), а с других, соседних тканей, в которых электрические поля этим органом создаются. В клиническом отношении это существенно упрощает саму процедуру регистрации, делая ее безопасной и несложной.
В электрографии для оценки функционального состояния органа по его электрической активности используется принцип эквивалентного генератора. Он состоит в том, что изучаемый орган, состоящий из множеств клеток, возбуждающихся в различные моменты времени, представляется моделью — единым эквивалентным генератором. Считается, что этот эквивалентный генератор находится внутри организма и создает на поверхности тела такое электрическое поле или распределение электрического потенциала, которое во все моменты времени как можно точнее должно совпадать с теми, что создает сам орган (это можно зарегистрировать в опыте). Очевидно, что чем выше эта точность, тем более адекватна реальному объекту выбранная модель.
Таким образом, потенциал электрического поля сердца можно представить в виде потенциала одного эквивалентного диполя.
Из формулы UАВ: UВС: UСА = pАВ: pВС: pСА (6) следует, что разность потенциалов между двумя точками диполя (на одинаковых расстояниях от него) пропорциональна проекции момента на линию, их соединяющую (рис. 7а,б).
Исследуя изменения разности потенциалов на поверхности человеческого тела, можно судить о проекциях дипольного момента сердца, следовательно, о биопотенциалах сердца.
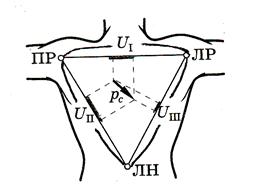
Рис. 7б (Треугольник Эйнтховена)
Эта идея стала одной из основных в модели Эйнтховена, основные постулаты которой можно сформулировать так:
— электрическое поле сердца представляется как электрическое поле точечного диполя с дипольным моментом, называемым интегральным электрическим вектором сердца (ИЭВС) (складывается из диполей разных частей сердца);
— ИЭВС находится в однородной проводящей среде, которой являются ткани организма;
— интегральный электрический вектор сердца меняется по величине и направлению. Его начало неподвижно и находится в атриовентрикулярном узле, а конец описывает сложную пространственную кривую, проекция которой на фронтальную плоскость образует за цикл сердечной деятельности (в норме) 3 петли: P, QRS и T. Изменение величины и направления вектора за 1 цикл сокращения сердца объясняется последовательностью распространения волн возбуждения по сердцу.
Эйнтховен предложил измерять разности потенциалов между двумя точками из трех, представляющими вершины равностороннего треугольника, в центре которого находится начало ИЭВС.
В практике электрокардиографии разности потенциалов измеряют между левой рукой (ЛР) и правой рукой (ПР) — I отведение, между левой ногой (ЛН) и правой рукой (ПР) - II отведение, между левой ногой (ЛН) и левой рукой (ЛР) - III отведение. Руки и ноги рассматриваются как проводники, отводящие потенциалы от вершин треугольника Эйнтховена, располагающихся у мест прикрепления конечностей к туловищу.
Связь между ИЭВС и разностью потенциалов показана на рис. 10.
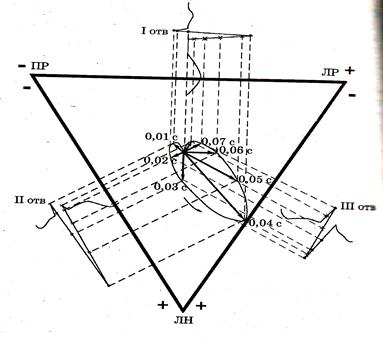
Рис.10
Электрокардиограмма – это график временной зависимости разности потенциалов в соответствующем отведении, а значит и временной зависимости проекции ИЭВС на линию отведения (рис. 11).
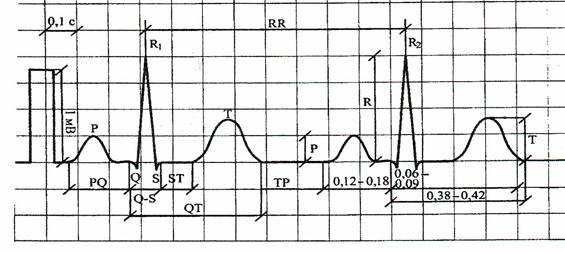
Рис. 11
Р – зубец отражает возбуждение предсердий;
P – Q интервал отсчитывается с начала зубца Р до начала зубца Q. Этот интервал отражает время, необходимое для деполяризации предсердий, проведения импульса через атриовентрикулярный узел, пучок Гиса и его ветви;
P – Q сегмент отсчитывается от конца зубца Р до начала зубца Q. Этот сегмент характеризует проводимость по атриовентрикулярному узлу, пучку Гиса, волокнам Пуркинье;
Q – зубец отражает начальный момент деполяризации межжелудочковой перегородки;
R – зубец отражает деполяризацию верхушки, задней и боковой стенок желудочков сердца;
S – зубец отражает процесс распространения волны возбуждения в базальных отделах межжелудочковой перегородки правого и левого желудочков;
QRS – комплекс отражает процесс деполяризации желудочков;
ST – сегмент - это отрезок от конца комплекса QRS до начала зубца Т. Он соответствует периоду угасания возбуждения желудочков и началу медленной реполяризации. Обычно ST – сегмент расположен на изолинии. У здорового человека смещение вверх не превышает 1 мм, вниз – 0.5 мм;
Т – зубец отражает процесс быстрой реполяризации желудочков;
Q – Т интервал соответствует электрической систоле желудочков;
U – зубец выявляется не всегда и считается, что он отражает реполяризацию волокон проводящей системы сердца. Лучше всего он выявляется в грудных отведениях;
Т – Р интервал – это отрезок ЭКГ от конца зубца Т, он соответствует состоянию покоя миокарда.
|
|
|
|
Дата добавления: 2014-01-03; Просмотров: 3707; Нарушение авторских прав?; Мы поможем в написании вашей работы!