КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Пути биосинтеза вторичных метаболитов
|
|
|
|
БИОХИМИЯ ВТОРИЧНОГО МЕТАБОЛИЗМА
Пути синтеза большинства вторичных метаболитов установлены достаточно хорошо. В настоящее время интенсивно изучается энзимология вторичного метаболизма. На основании имеющейся информации можно сформулировать некоторые закономерности биосинтеза этих соединений. Предшественниками синтеза служит относительно небольшое количество первичных метаболитов. Многие группы вторичных метаболитов могут синтезироваться несколькими путями. Часто этапы синтеза дублированы в разных компартментах клетки (например, пластиды — цитозоль). Синтез четко спланирован и обслуживается набором специальных ферментов, в большинстве случаев весьма специфичных.
Биосинтез алкалоидов. Образование этих веществ тесно связано с общим обменом азота клетки. Для большинства алкалоидов показано, что схемы их синтеза унифицированы, т. е. имеют сходную последовательность реакций. В процессе биосинтеза молекула аминокислоты практически полностью включается в структуру алкалоида. Синтез алкалоидов разных групп включает одинаковые типы реакций: декарбоксилирование, окислительное дезаминирование, альдольная конденсация, но для каждой группы алкалоидов эти реакции осуществляют «собственные» ферменты. На первом этапе синтеза происходит декарбоксилирование аминокислоты при участии соответствующей декарбоксилазы. Образовавшиеся биогенные амины подвергаются окислительному дезаминированию с участием аминооксидаз. Полученные в результате аминоальдегиды или аминокетоны в результате серии последовательных реакций образуют ключевые гетероциклические соединения. Затем происходит модификация базовой структуры с участием разнообразных реакций — гидроксилирования, метилирования и др. В формировании окончательной структуры алкалоида могут принимать участие дополнительные углеродные единицы, например ацетат (в виде ацетил-СоА) или монотерпеновая единица (для сложных индольных алкалоидов). В зависимости от сложности алкалоида его биосинтез включает от трех-четырех до десяти—пятнадцати реакций.
Для целого ряда алкалоидов не только установлена схема синтеза, но охарактеризованы и выделены ферменты. Оказалось, что некоторые ферменты синтеза не очень специфичны (в качестве субстратов могут использовать различные соединения), однако в цепочке синтеза обязательно присутствуют высокоспецифичные ферменты, которые используют только один субстрат (или ряд очень близких субстратов) и выполняют очень специфичную реакцию. Например, при синтезе изохинолинов гидроксилирование базовой структуры по каждому положению выполняют разные ферменты. По мере продвижения к заключительным этапам синтеза сродство ферментов к субстрату обычно повышается: например, для ряда ферментов синтеза бербериновых алкалоидов Кт составляет менее 1 мкМ. В качестве примера на рис. 9.12 представлена схема синтеза изохинолиновых алкалоидов.
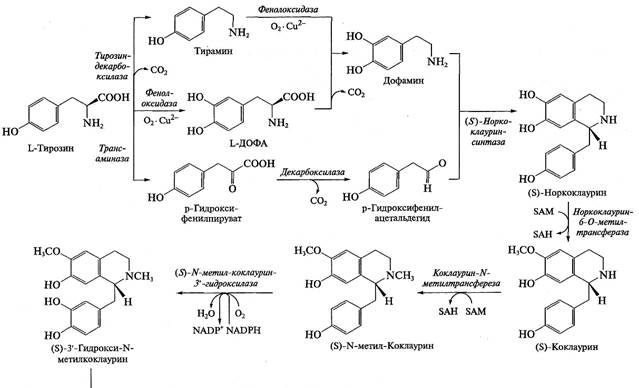
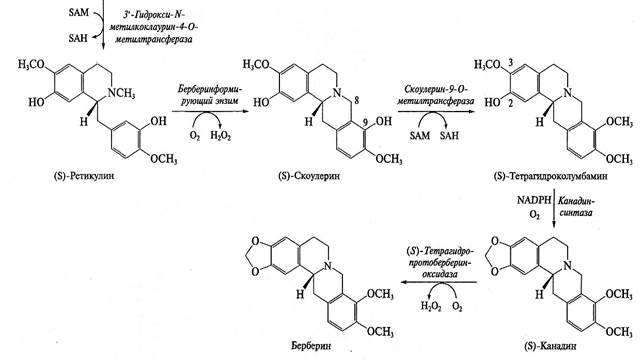
Рис. 9.12. Схема биосинтеза изохинолиновых алкалоидов
Биосинтез изопреноидов. Если при синтезе алкалоидов сходная цепочка превращений используется для различных исходных соединений (аминокислот), то синтез колоссального числа изопреноидов происходит из единственного предшественника — изопентенилдифосфата (ИПДФ). Под действием фермента изопентенилдифосфатизомеразы, которая сдвигает двойную связь, ИПДФ превращается в диметилаллилдифосфат (ДМАДФ). Далее ИПДФ присоединяется к ДМАДФ по двойной связи и образуется С10-соединение — геранилдифосфат. Он служит источником всех монотерпеноидов. Затем к геранилдифосфату присоединяется еще один ИПДФ и образуется С15-соединение фарнезилдифосфат — исходное вещество для синтеза сесквитерпеноидов. Далее фарнезилдифосфат может либо присоединить еще одну молекулу ИПДФ с образованием геранилгеранилдифосфата (С20-соединение — источник дитерпеноидов), либо димеризоваться с образованием сквалена (С30-соединение — исходное соединение для всех тритерпеноидов). Наконец, геранилгеранилдифосфат может димеризоваться с образованием фитоина — С40-соединения, источник тетратерпеноидов. Кроме того, к геранилгеранилдифосфату может последовательно присоединиться большое количество ИПДФ, формируя в конечном итоге полиизопреноиды — каучук и гуттаперчу. В результате описанных реакций образуется полный гомологический ряд С5-соединений разной длины. Далее эти алифатические молекулы могут «свернуться» в циклические структуры, причем количество циклов, их размер и типы сочленения могут быть самыми разными. На рис. 9.13 представлена общая схема синтеза изопреноидов.
Синтез базовых изопреноидных структур осуществляют всего два типа ферментов — пренилтрансферазы, которые «наращивают» длину изопреноидов, и циклазы, которые формируют соответствующий циклический скелет молекулы. При этом каждый тип структуры формирует специфическая циклаза. Так как типов циклических структур изопреноидов довольно много, то и количество циклаз должно быть внушительным. К настоящему времени их известно более ста. После формирования базовой структуры (или одновременно с этим), происходит ее модификация и «оснащение» функциональными группами.
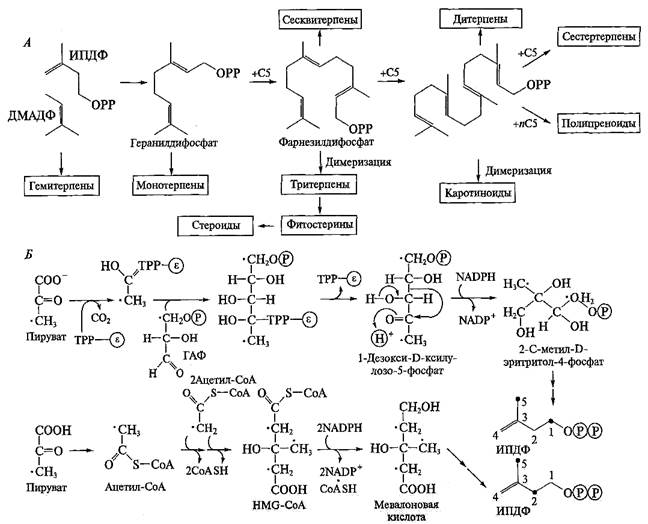
Рис. 9.13. Общая схема биосинтеза изопреноидов (Л) и два пути синтеза изопентенилдифосфата (Б) в растениях.
Точками показаны меченые атомы в исходных соединениях и в образовавшихся ИПДФ
Таким образом, биосинтез изопреноидов можно представить себе как своеобразный биохимический «моделист-конструктор». Вначале из унфицированых С5-модулей изготовляются гибкие линейные конструкции разной длины. Они представляют собой практически идеальный материал для «биохимического конструирования» и формирования множества вариантов циклических структур.
Долгое время считалось, что во всех организмах исходное соединение для образования изопреноидов — ИПДФ — формируется единственным способом, а именно из мевалоновой кислоты, которая в свою очередь синтезируется из трех молекул ацетил-СоА. Недавно было установлено, что разные организмы используют различные способы синтеза ИПДФ. В клетках животных и грибов все изопреноиды синтезируются по «классическому» мевалонатному пути. Целый ряд микроорганизмов, в том числе многие цианобактерии и зеленые водоросли, используют другой вариант образования ИПДФ. В этом случае его предшественником является 1-дезоксиксилулозо-5-фосфат, который синтезируется из пирувата и глицеральдегид-3-фосфата. Такой путь синтеза был назван «альтернативным», или «немевалонатным». Оказалось, что растения используют оба варианта образования изопреноидов: в цитозоле синтез идет по классическому пути, а в пластидах — по альтернативному (рис. 9.13). При этом возможно не только дублирование синтеза изопреноидов в разных ком-партментах клетки, но и разделение по типу синтезируемых структур. Тритерпеноиды (включая стероиды) синтезируются в цитозоле из мевалоната, тогда как дитерпеноиды (включая фитол хлорофилла) и тетратерпеноиды (прежде всего каротиноиды) — в пластидах по альтернативному пути. Моно- и сесквитерпены, вероятно, могут образовываться разными вариантами в зависимости от структуры молекулы и вида растения.
Биосинтез фенольных соединений. К настоящему времени известно два пути образования фенольных соединений — шикиматный (через шикимовую кислоту) и ацетатно-малонатный. Основной путь шикиматный, это практически единственный способ формирования ароматического кольца. В качестве исходных соединений для синтеза выступают фосфоенолпируват (ФЕП) и эритрозо-4-фосфат. При их конденсации возникает семиуглеродная кислота (2-кето-3-дезокси-7-фосфоарабогептановая), которая затем циклизуется в 5-дегидрохин-ную кислоту. Из дегидрохинной кислоты образуется шикимовая кислота, которая имеет шестичленное кольцо, одну двойную связь, и ее легко перевести в соединения ароматического ряда. Из шикимовой кислоты возможно образование оксибензойных кислот — n -оксибензойной, протокатеховой, галловой. Однако основной путь использования шикимовой кислоты — образование через префеновую кислоту ароматических аминокислот фенилаланина и тирозина. Фенилаланин (возможно, в ряде случаев и тирозин) — основной предшественник синтеза фенольных соединений. Дезаминирование фенилаланина осуществляет фермент фенилаланинаммиаклиаза (ФАЛ). В результате образуется коричная кислота, гидроксилирование которой приводит к образованию пара- кумаровой (оксикоричной) кислоты. После дополнительного гидроксилирования и последующего метилирования из нее образуются остальные оксикоричные кислоты.
Оксикоричные кислоты представляют центральное звено синтеза всех фенольных соединений клетки. Opтo -кумаровая кислота является предшественником кумаринов. После ряда реакций укорочения алифатической части молекулы образуются С6-С2- и С6-С1 – соединения — это второй путь образования оксибензойных кислот (первый — непосредственно из шикимовой кислоты). Оксикоричные кислоты могут образовывать различные конъюгаты, прежде всего с сахарами, однако основная масса оксикоричных кислот активируется путем взаимодействия с СоА. Два магистральных пути использования СоА-эфиров оксикоричных кислот — синтез лигнинов и синтез флавоноидов. Для синтеза лигнинов СоА-эфиры оксикоричных кислот восстанавливаются до спиртов, которые выступают в качестве мономеров синтеза. При синтезе флавоноидов СоА-производное оксикоричной кислоты взаимодействует с тремя молекулами малонил-СоА с образованием халкона. Реакцию катализирует фермент халконсинтаза. Образовавшийся халкон легко преобразуется в флаванон. Из флаванонов за счет реакций гидроксилирования, окисления — восстановления образуются другие группы флавоноидов. Затем может происходить модификация молекулы — гликозилирование, метоксилирование и др.
Ацетатно-малонатный путь синтеза фенольных соединений широко распространен у грибов, лишайников и микроорганизмов. У растений он является минорным. При синтезе соединений по этому пути ацетил-СоА карбоксилируется с образованием малонилацетил-СоА. Затем происходит каскад аналогичных реакций, в результате наращивается углеродная цепь и возникает
поли-β-кетометиленовая цепочка. Циклизация поликетидной цепи приводит к образованию различных фенольных соединений. Таким способом синтезируются флороглюцин и его производные, некоторые антрахиноны. В структуре флавоноидов кольцо В формируется по шикиматному пути (из оксикоричной кислоты), тогда как кольцо А — по ацетатно-малонатному.
В клетке работают два шикиматных пути синтеза флавоноидов — один в пластидах, другой в цитозоле. В этих компартментах находится полный набор изоферментов шикиматного пути, а также ферментов фенольного метаболизма, в том числе ФАЛ и халконсинтазы. Таким образом, в растительной клетке существует две параллельные цепочки синтеза фенольных соединений (аналогично изопреноидам).
Синтез минорных классов вторичных соединений. Образование этих веществ также изучено достаточно полно. Для многих азотсодержащих соединений исходными веществами являются аминокислоты. Например, синтез цианогенных гликозидов начинается с декарбоксилирования соответствующей аминокислоты, затем последовательно формируются альдоксим, нитрил и
α-гидроксинитрил. На последнем этапе синтеза образуется цианогенный гликозид за счет гликозилирования α-гидроксинитрила при помощи УДФ-глюкозы. Синтез обычно осуществляет комплекс ферментов: например, для дуррина этот комплекс состоит из четырех ферментов. Гены ферментов клонированы. Трансгенное по двум генам растение арабидопсиса приобрело способность к синтезу цианогенных гликозидов. Синтез беталаинов начинается от тирозина, который гидроксилируется и образуется диоксифенилаланин (ДОФА). ДОФА служит источником для двух фрагментов молекулы бетацианинов — беталамовой кислоты и цикло-ДОФА. Объединение этих двух соединений приводит к формированию бетацианинов. При синтезе бетаксантинов беталамовая кислота конденсируется с пролином. Серосодержащие вторичные метаболиты обычно синтезируются из серосодержащих аминокислот.
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 2909; Нарушение авторских прав?; Мы поможем в написании вашей работы!