КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Біосинтез жирних кислот
|
|
|
|
Біосинтез жирів.
Біосинтез жирів включає три основні процеси: біосинтез вищих жирних кислот, біосинтез гліцерину, сполучення складових частин у молекулу триацилгліцериду.
Біосинтез жирних кислот відбувається в основному у цитоплазмі з ацетил-КоА. Основними джерелами цієї сполуки є такі процеси: розщеплення жирних кислот, розщеплення кетонних амінокислот, розщеплення вуглеводів до піровиноградної кислоти, що здатна потім перетворюватися в ацетил-КоА.
Попередником жирних кислот є малоніл-КоА, а повний синтез насичених жирних кислот каталізується синтетазним комплексом, що складається з семи ферментів.
Синтетазна система, що каталізує біосинтез жирних кислот, знаходиться в розчинній фракції цитоплазми і відповідає за наступну сумарну реакцію при біосинтезі, наприклад, пальмітинової кислоти:
|
У цій реакції одна молекула ацетил-КоА конденсується з сімома молекулами малоніл-КоА з утворенням однієї молекули пальмітинової кислоти. Відновлення відбувається за рахунок НАДФ×Н2, що утворюються при фосфоглюконатному шляху перетворення глюкози.
[1]. Малоніл-КоА утворюється з цитоплазматичного ацетил-КоА та вуглекислого газу під дією ферменту ацетил-КоА-карбоксилази, коферментом якого є біотин:
 |
Фермент ацетил-КоА-карбоксилаза має регуляторні функції і тому лімітує весь процес синтезу жирних кислот.
[2]. Ацильні групи ацетил-КоА і малоніл-КоА переносяться на тіолові групи білка – ацилпереносника (АПБ), відповідно ацетилтрансацилазою і малонілтрансацилазою за схемою:
 | |||
 |
[3]. Наступним етапом є реакція конденсації, в якій ацетил-S-АПБ і малоніл-S-АПБ на поверхні ферменту реагують між собою з виділенням карбоксильної групи малоніл-S-АПБ:
 |
У результаті цієї реакції утворюється ацетоацетил-S-АПБ, а молекула СО2, яка виділилася, є той самий атом вуглецю, який приєднався із СО2 (див. реакцію ).
[4]. Ацетоацетил-S-АПБ відновлюється НАДФ×Н2 з утворенням стереоізомера D-b-оксибутирил-S-АПБ за допомогою ферменту b-кетоацил-АПБ-редуктази.
 |
[5]. Реакція дегідратації за допомогою ферменту еноїл-АПБ-дегідратази відбувається з утворенням транс-a,b- або D2,3 – ненасиченого ацильного АПБ-кротоніл-S-АПБ;
 |
[6]. Кротоніл-S-АПБ відновлюється потім кротоніл-АПБ-редуктазою до бутирил-S-АПБ. Донором водню, як і при першому відновленні, є НАДФ×Н2.
 |
Утворенням бутирил-S-АПБ завершується перший цикл, в якому молекула малоніл-S-АПБ приєднується до карбоксильного кінця ланцюга жирної кислоти. При цьому відщеплюється молекула АПБ, а кінцева карбоксильна група малоніл-S-АПБ відщеплюється у вигляді СО2.
Далі цикл реакцій повторюється. Допустимо, що йде синтез пальмітинової кислоти (С16). В цьому випадку утворенням бутирил-АПБ завершується лише перший з 7 циклів, в кожному з яких початком є приєднання молекули малоніл-АПБ до карбоксильного кінця ланцюга жирної кислоти, що росте. При цьому відщеплюється карбоксильна група малоніл-АПБ у вигляді СО2. Наприклад, бутиріл-АПБ, що утворився в першому циклі, взаємодіє з малоніл-АПБ:
 | |||
 | |||
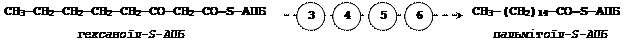 |
Завершується синтез жирної кислоти відщепленням HS-АПБ від ацил-АПБ під впливом ферменту деацилази.
 |
Отже, враховуючи, що на утворення однієї молекули малоніл-КоА з ацетіл-КоА витрачаються одна молекула АТФ і одна молекула СО2, яка потім відщеплюється, сумарне рівняння синтезу пальмітинової кислоти можна представити в такому вигляді:
 |
Швидкість біосинтезу жирних кислот визначається очевидно швидкістю утворення триацилгліцеридів і фосфоглідеридів тому, що вільні жирні кислоти є в клітинах у невеликих кількостях і не накопичуються.
У мітохондріях молекули насичених жирних кислот, що мають від 12 до 16 атомів вуглецю, подовжуються за рахунок послідовного приєднання ацетил-КоА.
Ненасичені жирні кислоти утворюються з насичених у клітинах за участю ферменту оксигенази.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 12277; Нарушение авторских прав?; Мы поможем в написании вашей работы!