КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Фосфорилирование глюкозы
|
|
|
|
Влияние инсулина на перемещение транспортёров глюкозы из цитоплазмы в плазматическую мембрану.
1 - связывание инсулина с рецептором; 2 - участок инсулинового рецептора, обращённый внутрь клетки, стимулирует перемещение транспортёров глюкозы; 3, 4 - транспортёры в составе содержащих их везикул перемещаются к плазматической мембране клетки, включаются в её состав и переносят глюкозу в клетку.
Известны различные нарушения в работе транспортёров глюкозы. Наследственный дефект этих белков может лежать в основе инсулинонезависимого сахарного диабета. Нарушения функции ГЛЮТ-4 возможны на следующих этапах:
· передача сигнала инсулина о перемещении этого транспортёра к мембране;
· перемещение транспортёра в цитоплазме;
· включение в состав мембраны;
· отшнуровывание от мембраны и т.д.
НАРУШЕНИЯ ПЕРЕВАРИВАНИЯ И ВСАСЫВАНИЯ УГЛЕВОДОВ
В основе патологии переваривания и всасывания углеводов могут быть причины двух типов:
· дефекты ферментов, участвующих в гидролизе углеводов в кишечнике;
· нарушение всасывания продуктов переваривания углеводов в клетки слизистой оболочки кишечника.
В обоих случаях возникают нерасщеплённые дисахариды или моносахариды. Эти невостребованные углеводы поступают в дистальные отделы кишечника, изменяя осмотическое давление содержимого кишечника. Кроме того, оставшиеся в просвете кишечника углеводы частично подвергаются ферментативному расщеплению микроорганизмами с образованием органических кислот и газов. Всё вместе приводит к притоку воды в кишечник, увеличению объёма кишечного содержимого, усилению перистальтики, спазмам и болям, а также метеоризму.
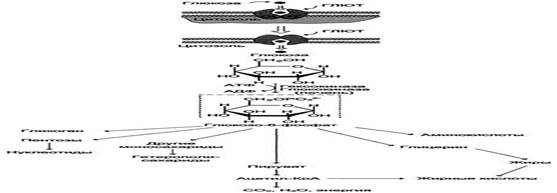
МЕТАБОЛИЗМ ГЛЮКОЗЫ В КЛЕТКЕ
После всасывания в кишечнике моносахариды поступают в воротную вену и далее преимущественно в печень. Поскольку в составе основных углеводов пищи преобладает глюкоза, её можно считать основным продуктом переваривания углеводов. Другие моносахариды, поступающие из кишечника в процессе метаболизма, могут превращаться в глюкозу или продукты её метаболизма. Часть глюкозы в печени депонируется в виде гликогена, а другая часть через общий кровоток доставляется и используется разными тканями и органами. При нормальном рационе питания концентрация глюкозы в крови поддерживается на уровне -3,3-5,5 ммоль/л. А в период пищеварения её концентрация может повышаться примерно 8 ммоль/л.
Метаболизм глюкозы в клетках всех тканей начинается с реакции фосфорилирования и превращения в глюкозо-6-фосфат (с использованием АТФ). Существуют два фермента, катализирующих фосфорилирование глюкозы: в печени и поджелудочной железе - фермент глюкокиназа, во всех других тканях – гексокиназа. Фосфорилирование глюкозы необратимая реакция, так как она протекает с использованием значительного количества энергии. Плазматическая мембрана клеток непроницаема для фосфорилированной глюкозы (нет соответствующих транспортных белков) и, следовательно, она уже не может из них выйти. Кроме того, фосфорилирование уменьшает концентрацию свободной глюкозы в цитоплазме. В результате создаются благоприятные условия для облегчённой диффузии глюкозы в клетки из крови.
Эти ферменты отличаются сродством к глюкозе. Гексокиназа имеет высокое сродство к глюкозе, т.е. этот фермент, в отличие от глюкокиназы, активен при низкой концентрации глюкозы в крови. Вследствие этого мозг, эритроциты и другие ткани могут использовать глюкозу при снижении ее концентрации в крови через 4-5 часов после еды и при голодании. Фермент гексокиназа может катализировать фосфорилирование не только D-глюкозы, но и других гексоз, хотя и с меньшей скоростью. Активность гексокиназы изменяется в зависимости от потребностей клетки в энергии. В качестве регуляторов выступают соотношение АТФ/АДФ и внутриклеточный уровень глюкозо-6-фосфата. При снижении расхода энергии в клетке повышается уровень АТФ (относительно АДФ) и глюкозо-6-фосфата. В этом случае активность гексокиназы снижается, и, следовательно, уменьшается скорость поступления глюкозы в клетку.
Фосфорилирование глюкозы в гепатоцитах в период пищеварения обеспечивается свойствами глюкокиназы. Активность глюкокиназы, в отличие от гексокиназы, не ингибируется глюкозо-6-фосфатом. Это обстоятельство обеспечивает повышение концентрации глюкозы в клетке в фосфорилированной форме, соответственно её уровню в крови. Глюкоза проникает в гепатоциты путём облегчённой диффузии при участии транспортёра ГЛЮТ-2 (независимого от инсулина). ГЛЮТ-2, так же, как глюкокиназа, имеет высокое сродство к глюкозе и способствует повышению скорости поступления глюкозы в гепатоциты в период пищеварения, т.е. ускоряет её фосфорилирование и дальнейшее использование для депонирования.
Хотя инсулин и не влияет на транспорт глюкозы, он усиливает приток глюкозы в гепатоциты в период пищеварения косвенным путём, индуцируя синтез глюкокиназы и ускоряя тем самым фосфорилирование глюкозы.
Преимущественное потребление глюкозы гепатоцитами, обусловленное свойствами глюкокиназы, предотвращает чрезмерное повышение её концентрации в крови в абсорбтивном периоде. Это, в свою очередь, снижает последствия протекания нежелательных реакций с участием глюкозы, например гликозилирования белков.
Дефосфорилирование глюкозо-6-фосфата
Превращение глюкозо-6-фосфата в глюкозу возможно в печени, почках и клетках эпителия кишечника. В клетках этих органов имеется фермент глюкозо-6-фосфатаза, катализирующая отщепление фосфатной группы гидролитическим путём:
Глюкозо-6-фосфат +Н2О → Глюкоза + Н3РО4
Образовавшаяся свободная глюкоза способна диффундировать из этих органов в кровь. В других органах и тканях глюкозо-6-фосфатазы нет, и поэтому дефосфорилирование глюкозо-6-фосфата невозможно. Пример подобного необратимого проникновения глюкозы в клетку - мышцы, где глюкозо-6-фосфат может использоваться только в метаболизме этой клетки.
Метаболизм глюкозо-6-фосфата
В зависимости от физиологического состояния организма и типа ткани глюкозо-6-фосфат может использоваться в клетке в различных превращениях, основными из которых являются: синтез гликогена, катаболизм с образованием СО2 и Н2О, синтез пентоз. Распад глюкозы до конечных продуктов служит источником энергии для организма. Вместе с тем в процессе метаболизма глюкозо-6-фосфата образуются промежуточные продукты, используемые в дальнейшем для синтеза аминокислот, нуклеотидов, глицерина и жирных кислот. Таким образом, глюкозо-6-фосфат - не только субстрат для окисления, но и строительный материал для синтеза новых соединений.
МЕТАБОЛИЗМ ГЛИКОГЕНА
Многие ткани синтезируют в качестве резервной формы глюкозы гликоген. Резервная роль гликогена обусловлена двумя важными свойствами: он осмотически неактивен и сильно ветвится, благодаря чему глюкоза быстро присоединяется к полимеру при биосинтезе и отщепляется при мобилизации. Синтез и распад гликогена обеспечивают постоянство концентрации глюкозы в крови и создают депо для её использования тканями по мере необходимости.
Строение и функции гликогена
Гликоген - разветвлённый полисахарид, в котором остатки глюкозы соединены в линейных участках α-1,4-гликозидной связью. В точках ветвления мономеры соединены α-1,6-гликозидными связями. Эти связи образуются примерно с каждым десятым остатком глюкозы, т.е. точки ветвления в гликогене встречаются примерно через каждые десять остатков глюкозы. Таким образом, в молекуле гликогена имеется только одна свободная аномерная ОН-группа и, следовательно, только один восстанавливающий (редуцирующий) конец.
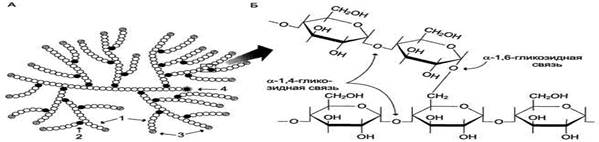
А. Строение молекулы гликогена: 1 - остатки глюкозы, соединённые α-1,4-гликозидной связью; 2 - остатки глюкозы, соединённые α-1,6-гликозидной связью; 3 - нередуцирующие концевые мономеры; 4 - редуцирующий концевой мономер.
Б. Строение отдельного фрагмента молекулы гликогена.
Гликоген хранится в цитозоле клетки в форме гранул диаметром 10-40 нм. С гранулами связаны и некоторые ферменты, участвующие в метаболизме гликогена, что облегчает их взаимодействие с субстратом. Разветвлённая структура гликогена обусловливает большое количество концевых мономеров, что способствует работе ферментов, отщепляющих или присоединяющих мономеры при распаде или синтезе гликогена, так как эти ферменты могут одновременно работать на нескольких ветвях молекулы. Гликоген депонируется главным образом в печени и скелетных мышцах.
После приёма пищи, богатой углеводами, запас гликогена в печени может составлять примерно 5% от её массы. В мышцах запасается около 1% гликогена, однако масса мышечной ткани значительно больше и поэтому общее количество гликогена в мышцах в 2 раза больше, чем в печени. Гликоген может синтезироваться во многих клетках, например в нейронах, макрофагах, клетках жировой ткани, но содержание его в этих тканях незначительно. В организме может содержаться до 450 г гликогена.
Распад гликогена печени служит в основном для поддержания уровня глюкозы в крови. Поэтому содержание гликогена в печени изменяется в зависимости от ритма питания. При длительном голодании оно снижается почти до нуля. Гликоген мышц служит резервом глюкозы - источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови.
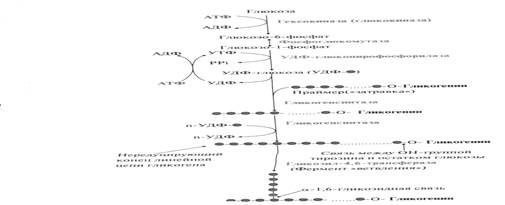
Синтез гликогена (гликогеногенез)
Гликоген синтезируется в период пищеварения (через 1-2 ч после приёма углеводной пищи). Следует отметить, что синтез гликогена из глюкозы требует затрат энергии.
Глюкоза активно поступает из крови в ткани и фосфорилируется, превращаясь в глюкозо-6-фосфат. Затем глюкозо-6-фосфат превращается фосфоглюкомутазой в глюкозо-1-фосфат, из которой под действием (УДФ)-глюкопирофосфорилазы и при участии (УТФ) образуется УДФ-глюкоза.
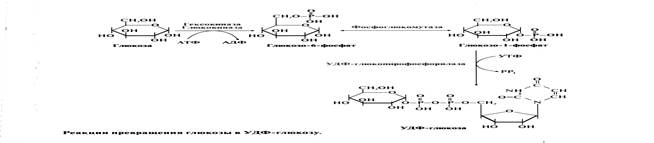
Но в силу обратимости реакции глюкозо-6-фосфат ↔ глюкозо-1-фосфат синтез гликогена из глюкозо-1-фосфата и его распад оказались бы также обратимыми и поэтому неконтролируемыми. Чтобы синтез гликогена был термодинамически необратимым, необходима дополнительная стадия образования уридиндифосфатглюкозы из УТФ и глюкозо-1-фосфата. Фермент, катализирующий эту реакцию, назван по обратной реакции: УДФ-глюкопирофосфорилаза. Однако в клетке обратная реакция не протекает, потому что образовавшийся в ходе прямой реакции пирофосфат очень быстро расщепляется пирофосфатазой на 2 молекулы фосфата. 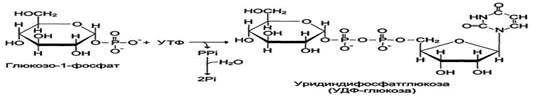
Образованная УДФ-глюкоза далее используется как донор остатка глюкозы при синтезе гликогена. Эту реакцию катализирует фермент гликогенсинтаза (глюкозилтрансфераза). Поскольку в данной реакции не используется АТФ, фермент называют синтазой, а не синтетазой. Фермент переносит остаток глюкозы на олигосахарид, состоящий из 6-10 остатков глюкозы и представляющий собой праймер (затравку), присоединяя молекулы глюкозы, α-1,4-гликозидными связями. Поскольку праймер редуцирующим концом соединен с ОН-группой остатка тирозина белка гликогенина, то гликогенсинтаза последовательно присоединяет глюкозу к нередуцирующему концу. Когда количество мономеров в синтезирующемся полисахариде достигает 11-12 моносахаридных остатков, фермент ветвления (гликозил-4,6-трансфераза) переносит фрагмент, содержащий 6-8 мономеров, то конца молекулы ближе к ее середине и присоединяет его α-1,6-гликозидной связью. В итоге образуется сильно разветвленный полисахарид.
Распад гликогена (гликогенолиз)
Распад гликогена или его мобилизация происходят в ответ на повышение потребности организма в глюкозе. Гликоген печени распадается в основном в интервалах между приёмами пищи, кроме того, этот процесс в печени и мышцах ускоряется во время физической работы.
Сначала фермент гликогенфосфорилаза расщепляет только α-1,4-гликозидные связи при участии фосфорной кислоты последовательно отщепляет остатки глюкозы от нередуцирующих концов молекулы гликогена и фосфорилирует их с образованием глюкозо-1-фосфата. Это приводит к укорочению ветвей.
Когда количество остатков глюкозы ветвях гликогена достигает 4, то фермент олигосахаридтрансфераза расщепляет α-1,4-гликозидную связь и переносит фрагмент, состоящий из 3 мономеров, к концу более длинной цепи.
Фермент α-1,6-гликозидаза гидролизует α-1,6-гликозидную связь в точке ветвления и отщепляет молекулу глюкозы. Таким образом, при мобилизации гликогена образуются глюкозо-1-фосфат и небольшое количество свободной глюкозы. Далее глюкозо-1-фосфат при участии фермента фосфоглюкомутазы превращается в глюкозо-6-фосфат.
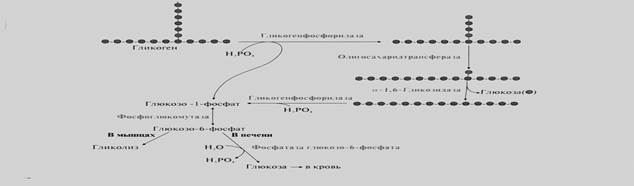
Мобилизация гликогена в печени и мышцах идет одинаково до образования глюкозо-6-фосфата. В печени под действием глюкозо-6-фосфатазы глюкозо-6-фосфат превращается в свободную глюкозу, которая поступает в кровь. Следовательно, мобилизация гликогена в печени обеспечивает сохранение нормального уровня глюкозы в крови и снабжение глюкозой других тканей. В мышцах нет фермента глюкозо-6-фосфатазы и глюкозо-6-фосфат используется самими мышцами для энергетических целей.
Биологическое значение обмена гликогена в печени и мышцах
Сравнение процессов синтеза и распада гликогена позволяет сделать следующие выводы:
· синтез и распад гликогена протекают по разным метаболическими путям;
· печень запасает глюкозу в виде гликогена не столько для собственных нужд, сколько для поддержания постоянной концентрации глюкозы в крови, и, следовательно, обеспечивает поступление глюкозы в другие ткани. Присутствие в печени глюкозо-6-фосфатазы обусловливает эту главную функцию печени в обмене гликогена;
· функция мышечного гликогена заключается в освобождении глюкозо-6-фосфата, потребляемого в самой мышце для окисления и использования энергии;
· при синтезе гликогена необходимы 1 моль АТФ и 1 моль УТФ;
· распад гликогена до глюкозо-6-фосфата не требует энергии;
· необратимость процессов синтеза и распада гликогена обеспечивается их регуляцией.
Нарушения обмена гликогена приводят к различным болезням. Они возникают при мутациях в генах, кодирующих ферменты, которые участвуют в обмене гликогена. При этих заболеваниях наблюдается накопление гранул гликогена в печени, мышцах и других тканей, приводящее к повреждению клеток.
РЕГУЛЯЦИЯ СИНТЕЗА И МЕТАБОЛИЗМА ГЛИКОГЕНА
Метаболизм гликогена в печени и мышцах зависит от потребностей организма в глюкозе как источнике энергии. В печени депонирование и мобилизацию гликогена регулируют гормоны инсулин, глюкагон и адреналин.
Инсулин и глюкагон – гормоны-антагонисты, их синтез и секреция зависят от концентрации глюкозы в крови. В норме концентрация глюкозы в крови соответствует 3,3-5,5 ммоль/л. Отношение концентрации инсулина к концентрации глюкагона в крови называют инсулин-глюкагоновым индексом.
Когда уровень глюкозы в крови повышается секреция инсулина увеличивается (инсулин-глюкагоновый индекс повышается). Инсулин способствует поступлению глюкозы в инсулинзависимые ткани, ускоряет использование глюкозы для синтеза гликогена в печени и мышцах.
Когда уровень глюкозы в крови снижается секреция инсулина снижается (инсулин-глюкагоновый индекс понижается). Глюкагон ускоряет мобилизацию гликогена в печени, вследствие чего увеличивается поступление глюкозы из печени в кровь.
Инсулин - синтезируется и секретируется в кровь р-клетками островков Лангерханса поджелудочной железы. β-клетки чувствительны к изменениям содержания глюкозы в крови и секретируют инсулин в ответ на повышение её содержания после приёма пищи. Транспортный белок (ГЛЮТ-2), обеспечивающий поступление глюкозы в β-клетки, отличается низким сродством к ней. Следовательно, этот белок транспортирует глюкозу в клетку поджелудочной железы лишь после того, как её содержание в крови будет выше нормального уровня (более 5,5 ммоль/л). В β-клетках глюкоза фосфорилируется глюкокиназой, скорость фосфорилирования глюкозы глюкокиназой в β-клетках прямо пропорциональна её концентрации в крови.
Синтез инсулина регулируется глюкозой. Глюкоза непосредственно участвует в регуляции экспрессии гена инсулина.
Глюкагон - вырабатываемый α-клетками поджелудочной железы в ответ на снижение уровня глюкозы в крови. По химической природе глюкагон - пептид.
Секреция инсулина и глюкагона также регулируется глюкозой, которая стимулирует секрецию инсулина из β-клеток и подавляет секрецию глюкагона из α-клеток. Кроме того, сам инсулин снижает секрецию глюкагона.
При интенсивной мышечной работе и стрессе в кровь из надпочечников секретируется адреналин. Он ускоряет мобилизацию гликогена в печени и мышцах, обеспечивая тем самым клетки разных тканей глюкозой.
Регуляция активности гликогенфосфорилазы и глигогенсинтазы
Действие этих гормонов в конечном счете сводится к изменению скорости реакций, катализируемых ключевыми ферментами метаболических путей обмена гликогена – гликогенсинтазой и гликогенфосфорилазой, активность которых регулируется аллостерически и фосфорилированием/ десфорилированием.
Гликогенфосфорилаза существует в 2 формах:
1) фосфорилированная - активная (форма а);
2) дефосфорилированная - неактивная (форма в).
Фосфорилирование осуществляется путём переноса фосфатного остатка с АТФ на гидроксильную группу одного из сериновых остатков фермента. Следствие этого - конформационные изменения молекулы фермента и его активация.
Взаимопревращения 2 форм гликогенфосфорилазы обеспечиваются действием ферментов киназы фосфорилазы и фосфопротеинфосфатазы (фермент, структурно связанный с молекулами гликогена). В свою очередь, активность киназы фосфорилазы и фосфопротеинфосфатазы также регулируется путём фосфорилирования и дефосфорилирования.
Активация киназы фосфорилазы происходит под действием протеинкиназы А - ПКА (цАМФ-зависимой). цАМФ сначала активирует протеинкиназу А, которая фосфорилирует киназу фосфорилазы, переводя её в активное состояние, а та, в свою очередь, фосфорилирует гликогенфосфорилазу. Синтез цАМФ стимулируется адреналином и глюкагоном.
Активация фосфопротеинфосфатазы происходит в результате реакции фосфорилирования, катализируемой специфической протеинкиназой, которая, в свою очередь, активируется инсулином посредством каскада реакций с участием других белков и ферментов. Активируемая инсулином протеинкиназа фосфорилирует и тем самым активирует фосфопротеинфосфатазу. Активная фосфопротеинфосфатаза дефосфорилирует и, следовательно, инактивирует киназу фосфорилазы и гликогенфосфорилазу.
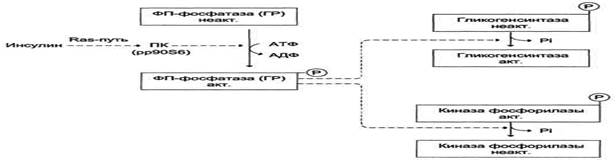
Влияние инсулина на активность гликогенсинтазы и киназы фосфорилазы. ФП-фосфатаза (ГР) - фосфопроте-инфосфатаза гранул гликогена. ПК (pp90S6) - протеинкиназа, активируемая инсулином.
Активность гликогенсинтазы также изменяется в результате фосфорилирования и дефосфорилирования. Однако есть существенные различия в регуляции гликогенфосфорилазы и гликогенсинтазы:
· фосфорилирование гликогенсинтазы катализирует ПК А и вызывает её инактивацию;
· дефосфорилирование гликогенсинтазы под действием фосфопротеинфосфатазы, наоборот, её активирует.
Регуляция метаболизма гликогена в печени
Повышение уровня глюкозы в крови стимулирует синтез и секрецию β-клетками поджелудочной железы гормона инсулина. Инсулин передает сигнал в клетку через мембранный каталитический рецептор - тирозиновую протеинкиназу. Взаимодействие рецептора с гормоном инициирует ряд последовательных реакций, приводящих к активации фосфопротеинфосфатазы гранул гликогена. Этот фермент дефосфорилирует гликогенсинтазу и гликогенфосфорилазу, в результате чего гликогенсинтаза активируется, а гликогенфосфорилаза становится неактивной.
Таким образом, в печени ускоряется синтез гликогена и тормозится его распад.
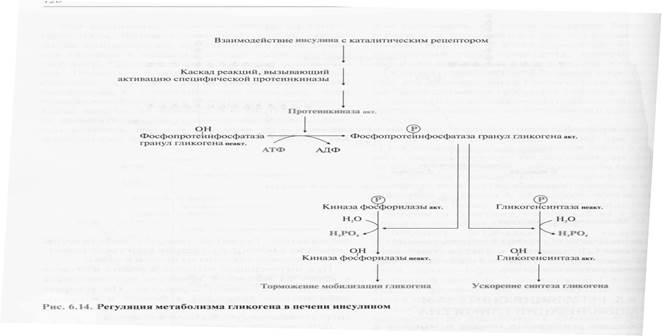
При голодании снижение уровня глюкозы в крови является сигналом для синтеза и секреции α-клетками поджелудочной железы глюкагона. Гормон передает сигнал в клетки через аденилатциклазную систему. Это приводит к активации проттеинкиназы А, которая фосфорилирует гликогенсинтазу и киназу фосфорилазы. В результате фосфорилирования гликогенсинтаза инактивируется и синтез гликогена тормозится, а киназа фосфорилазы становится активной и фосфорилирует гликогенфосфорилазу, которая становится активной. Активная гликогенфосфорилаза ускоряет мобилизацию гликогена в печени.
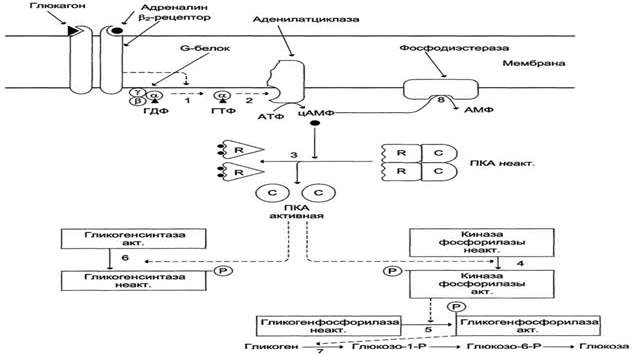
1 - глюкагон и адреналин взаимодействуют со специфическими мембранными рецепторами. Комплекс гормон-рецептор влияет на конформацию G-белка, вызывая диссоциацию его на протомеры и замену в α-субъединице ГДФ на ГТФ;
2 - α-субъединица, связанная с ГТФ, активирует аденилатциклазу, катализирующую синтез цАМФ из АТФ;
3 - в присутствии цАМФ протеинкиназа А обратимо диссоциирует, освобождая обладающие каталитической активностью субъединицы С;
4 - протеинкиназа А фосфорилирует и активирует киназу фосфорилазы;
5 - киназа фосфорилазы фосфорилирует гликогенфосфорилазу, переводя её в активную форму;
6 - протеинкиназа А фосфорилирует также гликогенсинтазу, переводя её в неактивное состояние;
7 - в результате ингибирования гликогенсинтазы и активации гликогенфосфорилазы гликоген включается в процесс распада;
8 - фосфодиэстераза катализирует распад цАМФ и тем самым прерывает действие гормонального сигнала. Комплекс α-субъединица-ГТФ затем распадается.
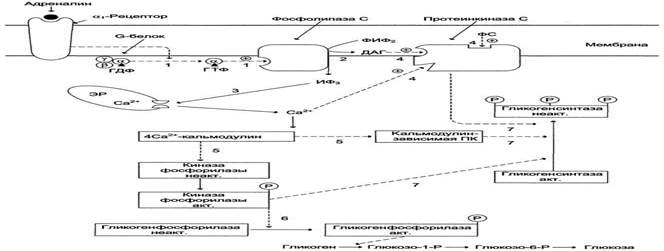
При интенсивной физическеой работе и стрессе в крови повышается концентрация а дреналина. В печени есть два типа мембранных рецепторов адреналина. Эффект адреналина в печени обусловлен фосфорилированием и активацией гликогенфосфорилазы. Адреналин имеет сходный с глюкагоном механизм действия. Но возможно включение и другой эффекторной системы передачи сигнала в клетку печени.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 8407; Нарушение авторских прав?; Мы поможем в написании вашей работы!