КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
NB! Глюконеогенез – механизм синтеза глюкозы
|
|
|
|
Запасы гликогена в печени ограничены и после 12-18 часового голодания они исчезают полностью. Многие клетки нуждаются в постоянном обеспечении глюкозой (эритроциты, нейроны, мышечные клетки в анаэробных условиях). Глюконеогенез является тем метаболическим путем, который решает данную проблему. Глюконеогенез – это метаболический путь превращения неуглеводных соединений в глюкозу. Многие соединения могут участвовать в этом процессе. Это и молочная кислота, и ПВК, и аминокислоты, распадающихся до пирувата (аланин, цистеин, глицин, серин, треонин и др.), и глицерин, и пропиононил-КоА, и субстраты цикла Кребса (оксалацетат и др., рис. 5.8).
Глюконеогенез представляет собой модификацию таких процессов, как гликолиз и цикл Кребса. Большая часть реакций гликолиза обратима. Исключение составляют три реакции, которые катализируют гексокиназа, фосфофруктокиназа-1 и пируваткиназа и для преодоления этих реакций используются специальные ферменты, которые назвали ключевыми реакциями глюконеогенеза. Данные ферменты сосредоточены в печени и корковом веществе почек. В таблице 5.2. приводятся названия ферментов, катализирующих необратимые реакции гликолиза и соответствующих им ключевых ферментов глюконеоегенеза.
Таблица 5.2. Ключевые ферменты гликолиза и гликонеогенеза
| Ферменты гликолиза | Ферменты глюконеогенеза |
| Гексокиназа | Глюкозо-6-фосфатаза |
| Фосфофруктокиназа-1 (ФФК-1) | Фруктозо-1,6-дифосфатаза |
| Пируваткиназа | 1. Пируваткарбоксилаза 2. Фосфоенолпируваткарбоксикиназа |
При совместной работе таких ферментов существует проблема т.н. “пустых” субстратных циклов. При условии катализа прямой и обратной реакции разными ферментами, продукт, получаемый в прямой реакции, становится субстратом другого фермента, который катализирует обратную реакцию, превращая продукт вновь в субстрат первого фермента. Возникает опасность “холостого” прокручивания субстратов реакции. Проблема решается организацией многоуровневой регуляции, включающей реципрокную аллостерическую регуляцию и ковалентную модификацию структуры ферментов.
Принято считать начальным этапом глюконеогенеза реакции, идущие в обход пируваткиназной реакции гликолиза. Пируваткиназа – объект влияния регуляторных систем(рис.5.9), управляющих скоростью гликолиза, поэтому в условиях благоприятствующих глюконеогенезу (голодание и др.) активность этого фермента следует затормозить. Этому способствует повышение количества аланина, который является аллостерическим ингибитором пируваткиназы и усиление секреции глюкагона. Последний стимулирует образование цАМФ в гепатоцитах, активирующей протеинкиназу А. Фосфорилирование пируваткиназы под влиянием протеинкиназы А вызывает переход ее в неактивное состояние. Торможение пируваткиназы благоприятствует включению глюконеогенеза. 

 .
. 
Рис.5.9. Регуляция активности пируваткиназы
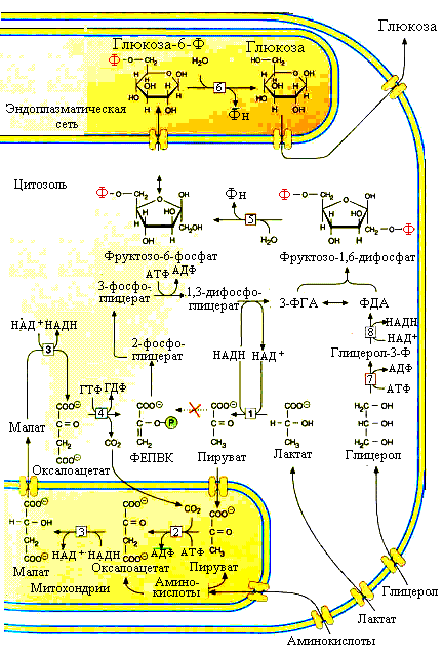
Рис.5.10. Основные субстраты и ферменты глюконеогенеза:
1–лактатдегидрогеназа; 2– пируваткарбоксилаза; 3–малатдегидрогеназа; 4–фосфоенолпируват карбоксикиназа; 5–фруктозо-1,6-дифосфатаза; 6– глюкозо-6-фосфатаза; 7–глицеролкиназа; 8–a-глицеролфосфатде гидрогеназа
Если превращение фосфоенолпирувата в ПВК, которое катализирует пируваткиназа, представляет одну химическую реакцию, то обратное превращение ПВК в фосфоенолпируват требует нескольких реакций. Первая реакция – это карбоксилирование пирувата. Реакция катализируется пируваткарбоксилазой и протекает с участием карбоксибиотина – активной форы СО2 в клетке. Продукт карбоксилирования – оксалоацетат занимает особое место в метаболизме митохондрий, где протекает данная реакция. Это важнейший субстрат цикла Кребса (см. ниже) и его выход из митохондрий затруднен. Для преодоления мембраны митохондрий оксалоацетат восстанавливается при помощи митохондриальной малатдегидрогеназы в легко приникающую через мебрану яблочную кислоту. Последняя, покинув митохондрии, в цитозоле окисляется вновь в оксалоацетат уже под влиянием цитозольной малатдегидрогеназы. Дальнейшее превращение оксалоацетата в ФЕПВК происходит в цитозоле клетки. Здесь при помощи фосфоенолпируваткарбоксикиназы окалоацетат декарбоксилируется с затратой энергии, высвобождаемой при гидролизе ГТФ и образуется ФЕПВК.
После образования ФЕПВК последующие реакции представляют обратимые реакции гликолиза. Из каждых двух образующихся 3-ФГА одна молекула при участии фосфотриозоизомеразы превращается в ФДА и обе триозы под влиянием альдолазы конденсируются в фруктозо-1,6-дифосфат. Некоторое количество ФДА образуется путем окисления глицеролфосфата, возникающего под влиянием глицеролкиназы из глицерола, поступающего в печень из жировой ткани. Это единственный субстрат из липидов, который участвует в глюконеогенезе. Превращение фруктозо-1,6-дифосфата во фруктозо-6-фосфат катализируется фруктозо-1,6-дифосфатазой-1. Затем вновь следует реакция, обратная гликолизу. Заключительная реакция глюконеогенеза катализируется ферментом глюкозо-6-фосфатазой, который катализирует гидролиз глюкозо-6-фосфатаи образующаяся свободная глюкоза может выходить из клетки.
Суммарная реакция синтеза молекулы глюкозы:
2 ПВК + 4 АТФ + 2 ГТФ + 2НАДН + 2H+ + 6H2O → Глюкоза + 2НАД+ + 4АДФ+ 2 ГДФ + 6 Фн +6H+
Таким образом, синтез одной молекулы глюкозы “обходится” клетке затратой шести макроэргов. 2 молекулы АТФ расходуются для активирования СО2, 2 молекулы ГТФ используются в фосфоенолпируваткарбоксикиназной реакции и 2 молекулы АТФ – для образования 1,3-дифосфоглицериновой кислоты.
Глюконеогенез активируется в клетках печени во время голодания, после продолжительных физических упражнений, при употреблении пищи, богатой белками при низком содержании в ней углеводов и т.д.
Интенсивность процесса зависит от количества субстратов, и активности, и количества ключевых ферментов гликолиза и глюконеогенеза.
Основными поставщиками субстратов для печени являются мышцы, эритроциты, жировая ткань. У последней довольно ограниченные возможности, поскольку только глицерол может использоваться для синтеза глюкозы, а это только около 6% от веса капельки жира.
Лактат, образующийся в результате работы мышц в анаэробных условиях или поступающий из эритроцитов, более значимый источник глюкозы. Наиболее важными источниками являются гликогенные аминокислоты, которые могут поступать с пищей, богатой белками или из мышц в условиях голодания.
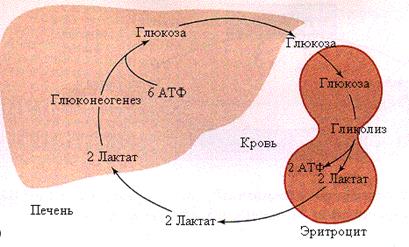
Рис. 5.11. Цикл Кори
Чтобы непрерывно снабжать глюкозой клетки, для которых она является основным источником энергии, но они не могут окислить ее полностью в силу отсутствия митохондрий (эритроциты) или из-за работы в анаэробных условиях, между печенью и этими клетками устанавливаются циклические процессы по обмену субстратами. Один из таких – цикл Кори: образующаяся в мышцах (эритроцитах) молочная кислота поступает в общий кровоток, захватывается печенью и используется ею в качестве субстрата глюконеогенеза; синтезируемая при этом глюкоза отдается в кровототок и метаболизируется мышцами или эритроцитами для получения энергии (рис. 5.11).
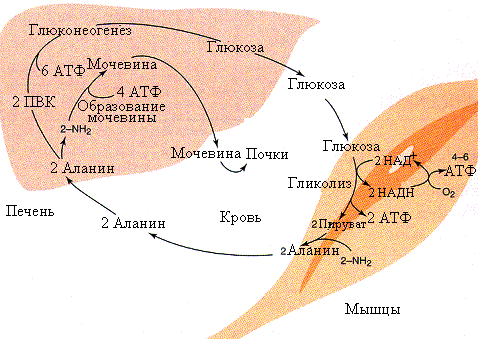
Рис.5.12.Аланиновый цикл
В отличие от цикла Кори, аланиновый цикл(рис.5.12) протекает при условии потребления периферическими тканями кислорода и требует митохондрий. При употреблении пищи богатой белами или при голодании происходит довольно активный обмен между печенью и мышцами аланином и глюкозой. Аланин из мышц передается клеткам печени, где он переаминируется и ПВК используется для синтеза глюкозы. По мере необходимости глюкоза поступает в мышцы и окисляется до ПВК, а затем, путем переаминирования, превращается в аланин который может вновь повторить этот цикл. Энергетически это более выгодный путь, чем цикл Кори.
|
|
|
|
|
Дата добавления: 2015-07-13; Просмотров: 1854; Нарушение авторских прав?; Мы поможем в написании вашей работы!