КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Біосинтез ліпідів
|
|
|
|
Біосинтез різних груп ліпідів має свої особливості.
Біосинтез жирів. Біосинтез жирів складається з трьох основних етапів: синтез гліцерину, вищих жирних кислот і сполучення їх в молекули тригліцеридів.
Синтез гліцерину. Близько 60% гліцерину утворюється з продукту проміжного обміну вуглеводів – діоксіацетонфосфата, решта поступає за рахунок тріоз пентозного циклу. Під впливом ферменту гліцерофосфатдегідрогенази діоксіацетонфосфат перетворюється в a-гліцерофосфат:
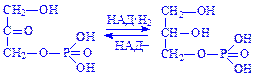
Частина гліцерину утворюється в результаті гідролізу гліцеридів клітин і міжклітинної рідини під впливом ліпаз:
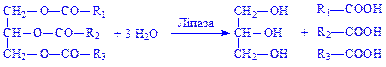
Далі гліцерин під впливом ферменту гліцеролкінази перетворюється в a-гліцерофосфат:
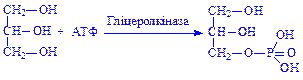
Фосфорилування гліцерину відбувається в більшості тканин і органів, окрім жирової тканини і слизової оболонки тонкої кишки, в яких фермент відсутній.
Біосинтез вищих жирних кислот. Головним компонентом, з якого утворюються вищі жирні кислоти, є ацетил-KoA. Він утворюється в процесі аеробного розпаду вуглеводів, b-окислення жирних кислот та ін.
Розрізняють два типи біосинтезу вищих жирних кислот: мітохондріальний і немітохондріальний. Перший з них протікає в основному в мітохондріях клітин багатьох органів (печінки, серця, мозку, нирок) і деяких тканин (жировій). Реакції каталізують ферменти синтетази, які локалізуються на мембранах органоїдів і здатні подовжувати молекули вже існуючих активованих залишків вищих жирних кислот за рахунок ацетил-КоА і НАД×Н2:
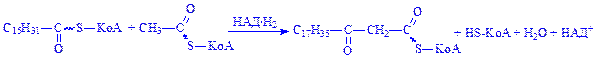
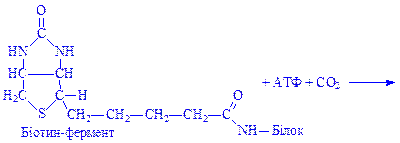
Другий тип біосинтезу вищих жирних кислот відбувається в гіалоплазмі різних клітин. У ньому беруть участь ферменти карбоксилази, НАДФ×Н2, АТФ і Mg2+. У складі молекул карбоксилази простетична група містить залишок вітаміну біотину, який здатний зв'язувати CO2.
На першому етапі біосинтезу відбувається вбудовування CO2 в молекулу біотин-ферменту (ацетил-КоА – карбоксилази) під впливом ферменту карбоксилази:

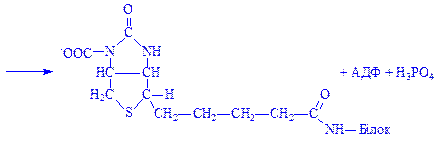
Потім CO2 переноситься на ацетил-КоА, що призводить до утворення малоніл-КоА:
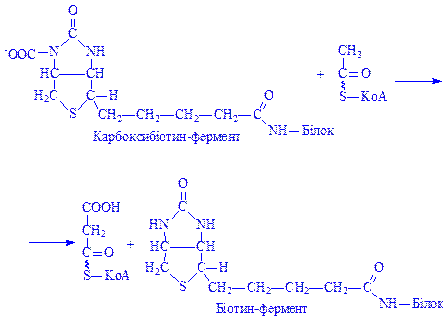
На наступному етапі йде поступове нарощування вуглеводневого радикала жирних кислот до потрібної величини за рахунок малоніл-КоА і ацетил-КоА з участю відповідних синтетаз. При цьому на кожному етапі ланцюг жирної кислоти подовжується на два вуглецеві атоми.
Якщо при біосинтезі утворюється пальмітинова кислота, то хімізм взаємодії ацетил-КоА і малоніл-КоА можна виразити так:
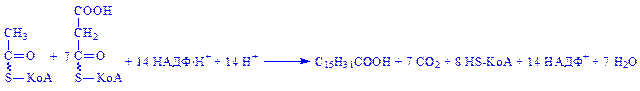
Останніми роками встановлений ще один шлях біосинтезу вищих жирних кислот – за допомогою білка ацилпереносника (АПБ), що містить простетичну групу, схожу з KoA. При цьому ацильні групи ацетил-КоА і малоніл-КоА переносяться на тіолові групи АПБ ферментами ацетил- і малонілтрансферазами. Koферментом є НАДФ×Н2, який утворюється в результаті функціонування пентозного циклу, окислення глутамінової кислоти, ін.
Біосинтез вищих жирних кислот з участю АПБ складається з ряду стадій:
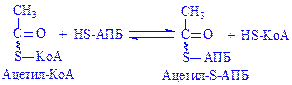 ;
; 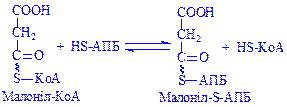 ;
;
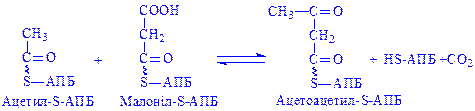 ;
;
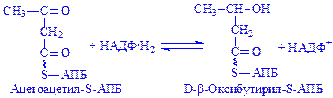 ;
; 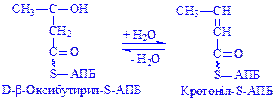 ;
; 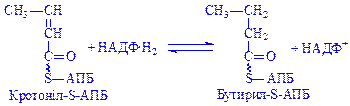 ;
;
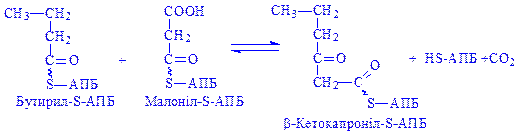
Далі відбувається нарощування вуглеводневого радикала жирної кислоти до величини, необхідної для біосинтезу потрібних клітині ліпідів.
Біосинтез тригліцеридів здійснюється в більшості тканин. Джерелом для синтезу є гліцерофосфат і ацил-KoA. Реакція протікає в декілька стадій. Спочатку утворюється фосфатидна кислота:

Реакцію каталізує мультиферментний комплекс – гліцерофосфатацилтрансфераза, яка локалізується в мікросомах.
Далі фосфатидна кислота під впливом ферменту фосфатидатфосфогідролази перетворюється в a-, b-дигліцерид – проміжний продукт біосинтезу жирів і фосфатидів:
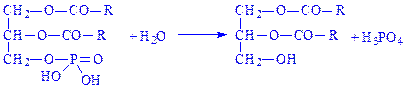
На останній стадії a-, b-дигліцерид під впливом того ж мультиферментного комплексу взаємодіє з ацил-КоА, утворюючи тригліцерид:

Біосинтез стеринів і стеридів. Стериди синтезуються з екзогенного і ендогенного холестерину та вищих жирних кислот. Найбільша кількість холестерину утворюється в тканинах печінки, головного мозку, наднирниках. Сировиною для біосинтезу служить оцтова кислота, оцтовий альдегід, ацетооцтова кислота, ацетон, ізовалеріанова кислота та ін. Будівельним блоком є ацетил-КоА. Біосинтез холестерину складається з ряду стадій, що включають конденсацію ацетилових одиниць в сквален, циклізацію сквалена в ланостерин, деметилювання ланостерина і утворення холестерину. В цих реакціях беруть участь багато ферментів, коферментів, гормонів, макроергічні сполуки (АТФ) та ін.
Розглянемо послідовність стадій біосинтезу холестерину.
1. Спочатку три молекули ацетил-КоА конденсуються у b-окси-b-метилглутарил-КоА:

2. b-Окси-b-метилглутарил-КоА під впливом ферменту редуктази перетворюється на мевалонову кислоту:
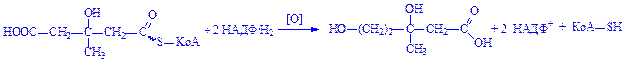
3. Під впливом ферменту кінази мевалонова кислота фосфорилується і перетворюється на пірофосфатмевалонову кислоту:

4. Під впливом ферменту декарбоксилази відбувається декарбоксилування пірофосфатмевалонової кислоти; утворюється ізопентилпірофосфат – „активний ізопрен”:
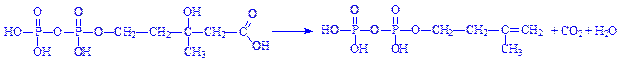
5. Під впливом ферменту ізомерази частина молекул ізопентилпірофосфата ізомеризується в диметилалілпірофосфат:
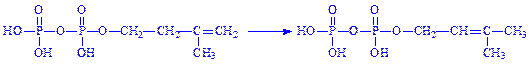
6. Молекули ізопентилпірофосфата і диметилалілпірофосфата конденсуються з утворенням геранілпірофосфата:
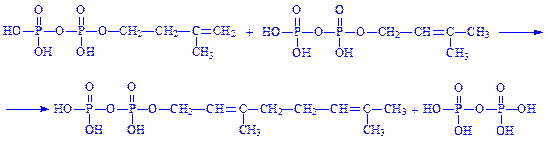
7. Геранілпірофосфат взаємодіє з ізопентилпірофосфатом з утворенням фарнезилпірофосфата:
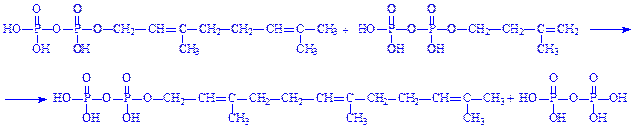
8. Дві молекули фарнезилпірофосфата конденсуються у вуглеводень сквален С30Н50, що містить шість подвійних зв'язків. Сквален в аеробних умовах циклізується, перетворюючись в ланостерин. Ланостерин деметилюється (втрачає три метильні групи), перетворюючись спочатку в зимостерин, потім – в десмостерин і, нарешті, в холестерин:
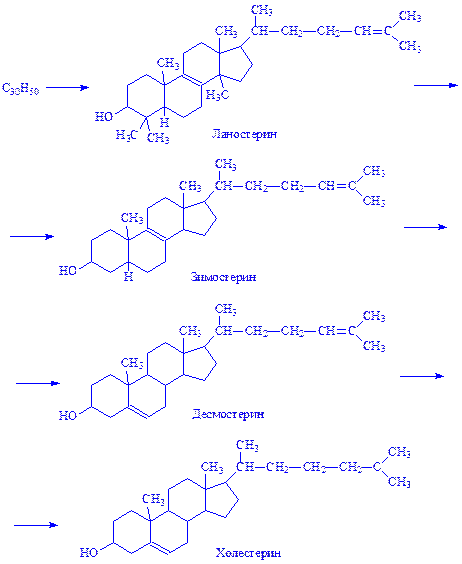
Невелика частина холестерину в тканинах синтезується з деяких амінокислот (лейцина). Після дезамінування і окислення утворюється ацетооцтова кислота – джерело b-окси-b-метилглутарил-KoA.
Холестерин характеризується високим ступенем обміну. Так, протягом доби 20% холестерину з міткою в бічному ланцюзі окислюється до CO2 і H2O. В окремих органах холестерин служить основою для біосинтезу молекул гормонів і біологічно важливих речовин (жовчних кислот).
Біосинтез фосфатидів. У тканинах і клітинах синтезуються окремі види гліцеро-, інозит- і сфінгозинфосфатидів. Перші стадії синтезу гліцерофосфатидів співпадають з такими при біосинтезі жирів (див. вище). Спочатку утворюються фосфатидні кислоти з фосфотріоз. Потім до них приєднуються вищі жирні кислоти у вигляді ацил-КоА. При утворенні молекули гліцерофосфатида один із залишків вищої жирної кислоти повинен належати ненасиченій жирній кислоті (олеїновій, лінолевій, ліноленовій), що поступає в організм у складі раціону. Під впливом фосфатаз фосфатидні кислоти гідролізуються, перетворюючись і a, b-дигліцериди. a, b-Дигліцериди взаємодіють з активованими азотистими основами, утворюючи молекули відповідних фосфатидів. Активація відбувається так:
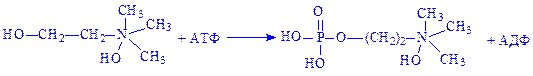
Азотиста основа фосфорилується. Холін під впливом ферменту холінкінази перетворюється на фосфорилхолін:
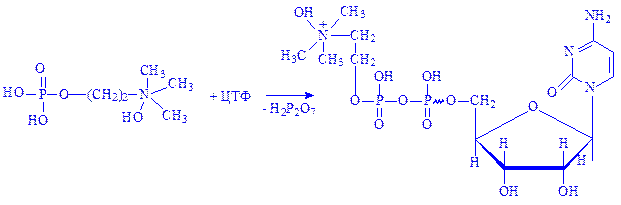
Фосфорилхолін під впливом ферменту холінфосфатцитидилилтрансферази взаємодіє з цитидинтрифосфатом (ЦТФ), перетворюючись в цитидиндифосфат-холін (ЦДФ-холін):
ЦДФ-холін під впливом ферменту холінфосфотрансферази вступає в реакцію з a, b-дигліцеридом, що призводить до утворення фосфатида і виділення цитидинмонофосфата (ЦМФ):
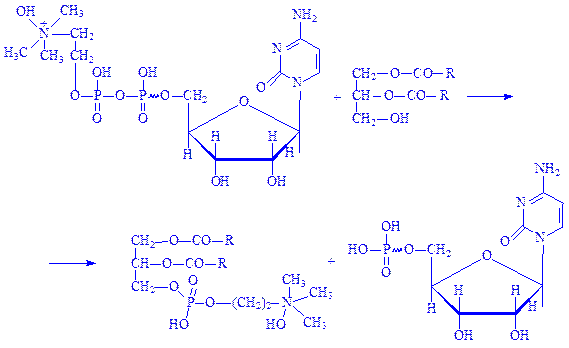
ЦМФ взаємодіє з АТФ під впливом ферменту ГТФ-аденілаткінази, перетворюючись в ЦТФ:
ЦМФ + 2АТФ ® ЦТФ + 2АДФ.
ЦТФ вступає в реакцію з новими порціями фосфатів азотистих основ, виконуючи функції специфічного каталізатора.
Біосинтез інших гліцерофосфатидів протікає аналогічно. В синтезі інозитфосфатидів центральне місце належить цитидиндифосфатдигліцериду, який взаємодіючи з молекулами інозиту, утворює моно-, ди- і триінозитфосфатиди.
Фосфатиди синтезуються у всіх клітинах тваринного організму. Найбільша кількість фосфатидів утворюється в печінці і тонкій кишці. Фосфатиди гепатоцитів запобігають печінковому ожирінню. Для фосфатидів характерний високий ступінь метаболізму.
Біосинтез гліколіпідів. Із всіх гліколіпідів найбільш детально вивчений біосинтез цереброзидів. У цьому процесі беруть участь багато ферментів, коферменти, іони Mn2+ та інші речовини. Реакції протікають в такій послідовності.
1. З пальмітил-КоА утворюється пальмітиновий альдегід:

2. Пальмітиновий альдегід конденсується з серином, що призводить до утворення дигідросфінгозина:
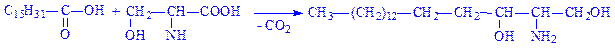
3. Дигідросфінгозин дегідрується, утворюючи сфінгозин:
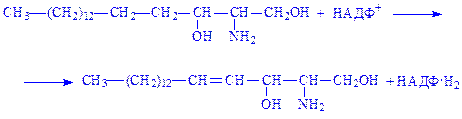
4. Відбувається ацилювання сфінгозина з утворенням цераміда:
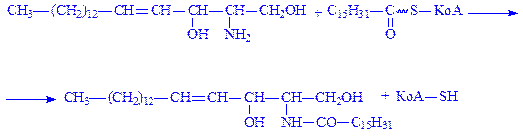
5. Церамід вступає в реакцію з УДФ-галактозою, що призводить до утворення цереброзида:
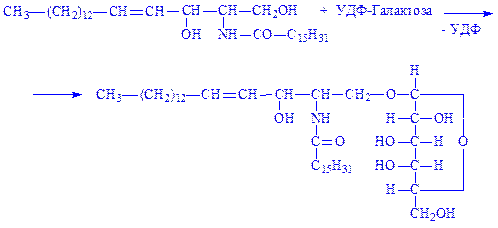
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 3308; Нарушение авторских прав?; Мы поможем в написании вашей работы!