КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Проміжний обмін
|
|
|
|
Продукти всмоктування білків через систему ворітної вени поступають у печінку. Амінокислоти, що залишилися в крові після проходження через печінку, з печінкової вени потрапляють у велике коло кровообігу і розносяться до окремих органів, тканин і клітин. Деяка частина амінокислот з міжклітинної рідини поступає в лімфатичну систему – лімфатичні капіляри ворсинок, підепітеліальну і підслизову сітку судин тонкої кишки, брижові вузли, грудну лімфатичну протоку і краніальну порожнисту вену.
Основна маса амінокислот витрачається на біосинтез білків, частина – на біосинтез біологічно активних речовин (небілкових гормонів, пептидів, амінів та ін.), частина, дезамінуючись, використовується як енергетична сировина і матеріал для біосинтезу ліпідів, вуглеводів, нуклеїнових кислот та ін.
Біосинтез білків. Проблема біосинтезу білка є однією з основних проблем біохімії. Вона має важливе теоретичне і практичне значення, тісно пов'язана з найактуальнішими питаннями сучасної біологічної науки: виясненням законів спадковості і мінливості, керуванням ростом і розвитком організмів, розкриттям причин виникнення і розробкою методів профілактики та лікування багатьох спадкових захворювань тощо.
Біосинтез білка протікає у всіх органах, тканинах і клітинах. Найбільша кількість білка синтезується в печінці. Синтез білка здійснюють рибосоми. За хімічною природою рибосоми – нуклеопротеїди, що складаються з РНК (50 – 65%) і білків (35 – 50%). У процесі біосинтезу білка рибосоми здійснюють функції: 1) специфічного скріплення і утримання компонентів білок-синтезуючої системи; 2) каталітичної підстанції (при утворенні пептидного зв'язку і гідролізі ГТФ); 3) транслокатора (при механічному переміщенні і- і тРНК).
Рибосоми утворюються самочинно із заздалегідь синтезованих РНК і білків. Вони є складовими частинами гранулярної ендоплазматичної сітки, яка є своєрідною транспортною системою клітини, де відбувається біосинтез і переміщення синтезованих молекул білка.
Рибосоми в клітині знаходяться у вигляді скупчень від 3 до 100 одиниць – полісом (полірибосом, ергосом). Рибосоми сполучені між собою своєрідною ниткою, видимою під електронним мікроскопом, – іРНК (рис. 2). Кожна рибосома здатна синтезувати самостійно один поліпептидний ланцюг, група – декілька таких ланцюгів і молекулу білка. Прикладом великої полірибосомної системи є полісоми м'язової тканини, які синтезують міозин. Полісома складається з 60 – 100 рибосом і здійснює біосинтез молекули білка, який складається з 1800 амінокислотних залишків.
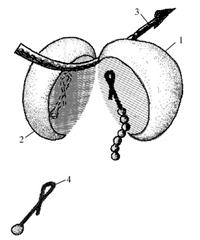
Рис 2. Схема будови рибосоми:
1 – 30S субодиниця; 2 – 50S субодиниця; 3 – іРНК; 4 – аміноацил-тРНК-фермент
Відомо, що в організмах людини і тварин синтезуються мільйони різних білків. Вони відрізняються один від одного, насамперед, хімічною природою та послідовністю розташування залишків амінокислот у поліпептидних ланцюгах, тобто первинною структурою. Інформація про те, яким повинен бути білок, закладена в ДНК у вигляді певної послідовності нуклеотидних залишків у полінуклеотидному ланцюгу.
Оскільки ДНК знаходиться в ядрі, а біосинтез білка відбувається на рибосомах, то ДНК передає інформацію щодо процесу синтезу білка через іРНК, яка синтезується на певній ділянці (гені) одного з нуклеотидних ланцюгів ДНК.
В основі передачі інформації лежить принцип комплементарності. У синтезованій іРНК послідовність нуклеотидів відповідає послідовності нуклеотидів в одному з полінуклеотидних ланцюгів ДНК. Відмінність полягає лише в тому, що замість тимідинового нуклеотиду в іРНК міститься уридиновий нуклеотид. Процес копіювання даної інформації з ДНК на іРНК називається транскрипцією.
Далі іРНК, діставши інформацію від ДНК, виходить з ядра і переміщується до рибосом. На рибосомах іРНК реалізує цю інформацію в процесі синтезу білка. Іншими словами, на іРНК як на матриці відбувається синтез білка, первинна структура якого визначається інформацією, що іРНК отримала від ДНК. Процес передачі інформації з іРНК, яка закодована в певній послідовності нуклеотидів в її молекулі, на процес розміщення залишків амінокислот у білкових молекулах називається трансляцією.
Отже, передачу інформації від ДНК на синтез білка можна подати схемою:
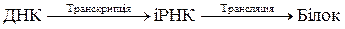 .
.
Для синтезу білків використовуються активовані форми амінокислот, які перебувають у зв'язаному стані з відповідними тРНК. Останні переносять їх до місця біосинтезу білка – рибосом. Процес сполучення амінокислот із „своїми” тРНК за участю ферменту аміноацил-тРНК – синтетази називають рекогніцією (від англ. recognice – пізнавати).
Згідно з сучасними уявленнями, біосинтез білків включає ряд складних біохімічних процесів, в яких беруть участь нуклеїнові кислоти, різні ферментні системи, іони металів тощо. Він поділяється на три основні стадії: транскрипцію, рекогніцію і трансляцію. Стадія транскрипції детально описана в темі „ обмін нуклеїнових кислот”, стадії рекогніції і трансляції описані нижче.
Рекогніція. Активація амінокислот відбувається у цитоплазмі за участю високоенергетичної сполуки – АТФ і ферментів – аміноацил-тРНК-синтетаз (аміноацил-синтетаз). Для кожної амінокислоти в клітині є специфічні ферменти, які називаються кодазами. Для виявлення ними максимальної активності необхідні, іони магнію. Дещо меншу активуючу дію мають іони інших двовалентних металів, зокрема марганцю, кобальту і кальцію. Реакцію активації амінокислот схематично можна подати так:
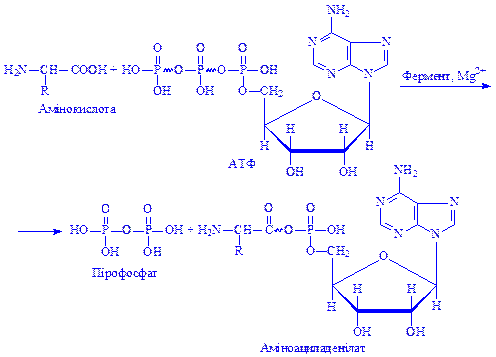
Ця реакція відбувається на поверхні ферменту, що каталізує її, і утворений аміноациладенілат не переходить у розчин, а залишається в комплексі з ферментом. У молекулі аміноациладенілату залишок амінокислоти сполучається з залишком АМФ макроергічним зв'язком, який посилює реакційну здатність амінокислоти.
На наступному етапі комплекс аміноациладенілату з ферментом взаємодіє з тРНК, специфічною для кожної амінокислоти. При цьому аміноацильна група з аміноациладенілату переходить до тРНК з утворенням нового комплексу – аміноацил-тРНК і виділяються АМФ та фермент:
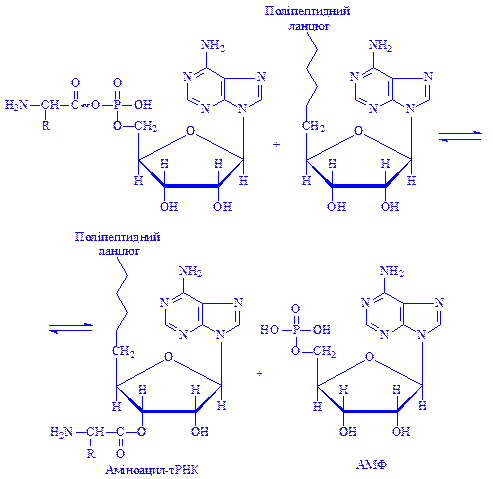
Залишок амінокислоти приєднується до третього атома вуглецю рибози кінцевого нуклеотиду тРНК, макроергічний зв'язок зберігається. Реакцію каталізує той самий фермент, що й реакцію активації амінокислоти – аміноацил-тРНК-синтетаза. У молекулі даного ферменту є дві специфічні ділянки, завдяки яким він здатний „впізнавати”, з однієї сторони, „свою” амінокислоту, а з другої – „свою” тРНК.
Деякі аміноацилсинтетази (наприклад, валінова, лейцинова, ізолейцинова) побудовані з одного поліпептидного ланцюга, інші – з двох, чотирьох і більшої кількості поліпептидних ланцюгів (наприклад, серинова аміноацилсинтетаза складається з двох субодиниць, метіонінова – з чотирьох). Молекулярна маса кожної субодиниці становить 45000. Виявлено аміноацилсинтетази, побудовані з двох і більшої кількості різних субодиниць. Наприклад, гліцинова аміноацилсинтетаза складається з чотирьох субодиниць, дві з яких мають молекулярну масу по 33000, а дві інші – по 80000.
Аміноацилсинтетази виявляють високу специфічність. Для кожної з 20 амінокислот, що входять до складу білків, є своя, і притому лише одна аміноацилсинтетаза. Водночас кожна з них „впізнає” всі тРНК, специфічні для однієї амінокислоти.
Трансляція. Трансляція включає три основні етапи – ініціацію, елонгацію і термінацію.
Ініціація синтезу поліпептидного ланцюга – це утворення ініціюючого комплексу і формування функціонально активної рибосоми. Розміри рибосом характеризують константами седиментації (виражають одиницями Сведберга S). Чим більші розміри мають часточки, тим швидше вони осідають під час ультрацентрифугування і тим вища константа їх седиментації.
Для прокаріот характерні рибосоми, константа седиментації яких становить 70S, а молекулярна маса 3×106. В еукаріотів рибосоми дещо більші. Константа їх седиментації досягає 80S, а молекулярна маса 4,5×106.
Кожна рибосома складається з двох субодиниць – малої і великої. У прокаріот 70S рибосоми побудовані з малої (30S) і великої (50S) субодиниць. Мала субодиниця містить молекулу 16S-PHK і 21 молекулу білків з різною молекулярною масою. 16S-РНК виконує в субодиниці структурну роль, вона необхідна для контакту рибосом з іРНК. Для контакту з іРНК у цій самій субодиниці є зона, в якій розміщена спеціальна акцепторна ділянка (Аср-ділянка) для зв'язування тРНК, яка доставляє активовані амінокислоти.
50S субодиниця містить дві молекули PHK: 23S-РНК, 5S-РНК і 34 молекули різних білків. 23S-РНК виконує структурну роль, а 5S-РНК необхідна для взаємодії субодиниці з тРНК. Отже, iРНК сполучається як з малою, так і з великою субодиницею рибосоми.
На 50S субодиниці рибосоми є дві ділянки – пептидильна, або П-ділянка, і аміноацильна, або А-ділянка; П-ділянка призначена для розміщення поліпептидного ланцюга, що синтезується, а в А-ділянку поступають аміноацил-тРНК.
Рибосоми еукаріот 80S побудовані з малої (40S) і великої (60S) субодиниць. Мала субодиниця містить молекулу 18S-РНК і близько 30 молекул різних білків. До складу великої субодиниці входять молекули 28S-PHK i 5S-РНК, а також близько 50 молекул різних білків. На субодиницях рибосом еукаріот також є система зон і ділянок для контакту і зв'язування iРНК, тРНК та ряду інших компонентів, необхідних для синтезу білка.
За певних умов рибосоми можуть розщеплюватись на субодиниці, тобто дисоціювати, а потім знову сполучатися – асоціювати, що має велике значення в процесі синтезу білка.
Для ініціації синтезу білка крім рибосом і рибонуклеїнових кислот важливим є наявність формілметіоніл-тРНК (тРНКфмет), факторів ініціації (1F-1, 1F-2, 1F-3), ГТФ, іонів магнію тощо.
Формілметіонін утворюється після приєднання метіоніну до тРНК за участю 10-формілтетрагідрофолієвої кислоти і специфічного ферменту – трансформілази. Така тРНК за будовою відрізняється від звичайної метіонінової тРНК. Обидві метіонін-тРНК (формілметіоніл-тРНК і метіоніл-тРНК) „впізнають” на іРНК триплет (кодон) АУГ. Однак даний триплет кодує формілметіонін лише тоді, коли він є початковим (ініціюючим), тобто коли з нього починається синтез поліпептидного ланцюга молекули білка. Якщо даний триплет розміщений не в тому місці іРНК, з якого починається синтез білкової молекули, а далі, всередині полінуклеотидного ланцюга іРНК, то він кодує звичайний метіонін.
Основна роль формільного залишку в метіоніні полягає в тому, що він, зв'язуючи аміногрупу, зумовлює взаємодію карбоксильної групи першої амінокислоти з аміногрупою наступної амінокислоти, тобто відбувається синтез поліпептидного ланцюга в напрямку NH2 ® СООН. Після закінчення синтезу поліпептидного ланцюга від нього може відщеплюватись формільна група або повністю формілметіонін.
Перед початком процесу ініціації рибосома розщеплюється на субодиниці. Ініціація розпочинається з утворення двох комплексів (рис. 3). Перший подвійний комплекс 1F-3-30S утворюється внаслідок взаємодії меншої субодиниці рибосоми 30S з фактором ініціації 1F-3 (І). Другий трикомпонентний 1F-2×ГТФ×тРНКфмет комплекс утворюється внаслідок взаємодії формілметіонін-тРНК з фактором ініціації 1F-2 і ГТФ (ІІ). Далі ці два комплекси сполучаються в один, який має склад 1F-3-30S-1F-2×ГТФ×тРНКфмет (ІІІ). На наступному етапі цей комплекс за участю фактора 1F-1 взаємодіє з іРНК, утворюючи ініціюючий комплекс (IV). Останній взаємодіє з 50S субодиницею рибосоми, що супроводжується вивільненням факторів ініціації і розщепленням ГТФ на ГДФ і H3PO4 (V). За цих умов утворюється функціонально активна 70S рибосома (30S×50S×PHK×тРНКфмет). Механізм ініціації синтезу поліпептидного ланцюга в еукаріот на 80S рибосомах в основному подібний до механізму цього процесу в прокаріот, проте має і специфічні особливості, зокрема число факторів ініціації в еукаріот значно більше, ніж у прокаріот, що, очевидно вказує на їх регулюючу дію.
Елонгація (ріст) поліпептидного ланцюга у момент закінчення ініціації формілметіонін-тРНК розміщена в П-ділянці рибосоми, а А-ділянка вільна і може приймати наступну аміноацил-тРНК.
Перший етап елонгації полягає в доставці аміноацил-тРНК до рибосоми і сполученні її з відповідним кодоном іРНК, розміщеним поряд з початковим (ініціюючим) кодоном АУГ. У цьому процесі беруть участь ГТФ і фактори елонгації EF–Tu, EF–Ts і EF–G (рис. 4). Спочатку перший фактор елонгації взаємодіє з ГТФ і аміноацил-тРНК з утворенням потрійного комплексу EF-Tu×ГТФ×аміноацил-тРНК. Далі цей комплекс доставляється в А-ділянку рибосоми, де й відбувається його дисоціація. Аміноацил-тРНК залишається зв'язаною з рибосомою, а ГТФ гідролізує до ГДФ і відщеплюється від рибосоми у вигляді ГДФ-EF-Tu. Під впливом другого фактора елонгації (EF–Ts) цей комплекс розкладається й обидва фактори знову включаються в доставку наступної молекули аміноацил-тРНК в А-ділянку рибосоми. Джерелом енергії служить нова молекула ГТФ.
На другому етапі елонгації формується пептидний зв'язок в А-ділянці, де розміщується доставлена відповідна аміноацил-тРНК. Сюди з П-ділянки переміщується формілметіонін-тРНК і утворюється перший пептидний зв'язок за рахунок етерифікованої карбоксильної групи формілметіонін-тРНК і аміногрупи, доставленої аміноацил-тРНК. При цьому утворюється пептидил-тРНК:
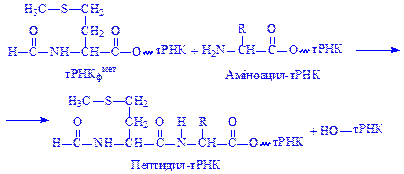
Процес утворення пептидного зв'язку каталізує фермент пептидил-трансфераза.
Вивільнена тРНК утримується в П-ділянці, а пептидил-тРНК в А-ділянці рибосоми.
На третьому етапі відбувається переміщення (транслокація) пептидил-тРНК з А-ділянки в П-ділянку. Цей процес проходить за рахунок енергії гідролізу другої молекули ГТФ. Реакцію каталізує білковий фактор EF-G, який часто називають рибосомо-залежною гуанозинтрифосфатазою.
Під час транслокації з П-ділянки видаляється вільна тРНК, а іРНК переміщується по рибосомі на довжину одного кодону. За цих умов А-ділянка вивільнюється і може приймати нову амінокислоту з тРНК.
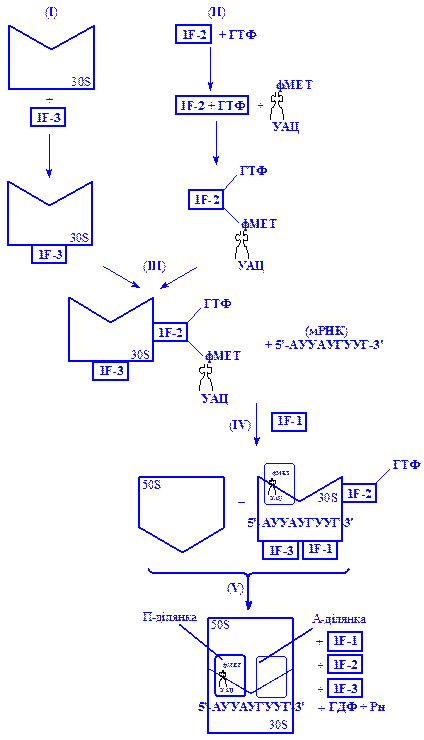
Рис. 3. Схема ініціації білкового синтезу (утворення ініціюючого
комплексу і формування активної рибосоми)
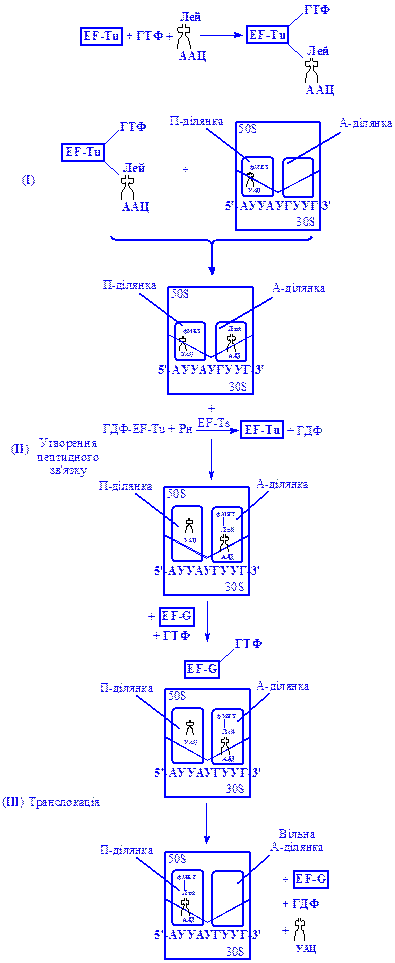
Рис 4. Основні етапи росту пептидного ланцюга
Термінація (закінчення синтезу) поліпептидного ланцюга. Термінація поліпептиду розпочинається при появі певних сигналів, якими є триплети УАА, УАГ, УГА. Ці триплети часто називають беззмістовними, термінаторними або нонсенс-триплетами. У процесі термінації беруть участь фактори RF1 і RF2, які каталізують відщеплення синтезованих поліпептидних ланцюгів від рибосоми. Фактор RF1 реагує на появу УАГ і УАА, фактор RF2 – на УАА і УГА.
В еукаріот термінація проходить за участю лише одного фактора – RF, для дії якого необхідна ГТФ.
Процес термінації відбувається в кілька етапів. Спочатку поліпептидний ланцюг з А-ділянки рибосоми, де утворюється останній пептидний зв'язок, переміщується в П-ділянку. Далі розривається складноефірний зв'язок між С-кінцевою амінокислотою та її тРНК і білок виходить із рибосоми. Комплекс рибосома×іРНК×тРНК розкладається, рибосома дисоціює на субодиниці. Вивільнені рибонуклеїнові кислоти, очевидно, можуть використовуватись у наступних циклах синтезу білка. Даний процес досить складний і повністю ще не вивчений.
Обчислено, що синтез типового білка, побудованого з ~150 – 200 залишків амінокислот на рибосомах, триває 1 – 3 xв. Отже, кожний цикл роботи рибосоми, що забезпечує подовження поліпептидного ланцюга білка на один залишок амінокислоти, триває частки секунди. З'ясування основних етапів синтезу білка є значним досягненням біології на молекулярному рівні. Уже експериментально встановлена генетична роль нуклеїнових кислот, розкрита суть генетичного коду, основаного на молекулярній структурі ДНК, і цим самим конкретизована природа мутацій – основа еволюції і мінливості живих організмів.
У гіалоплазмі з поліпептидних ланцюгів утворюються відповідні прості і складні білки. Формується вторинна, третинна і у ряді випадків четвертна структура білкової молекули.
Оновлення білків в організмі. Білки знаходяться в динамічному стані і піддаються постійним процесам синтезу і розпаду. У ході життєдіяльності білки поступово „зношуються” – руйнуються їх четвертна, третинна, вторинна і первинна структури. Інактивуются білкові функціональні групи і руйнуються зв'язки в білковій молекулі. Виникає необхідність в заміні „зношених білкових молекул” новими.
В залежності, від ступеня пошкодження білкової молекули походить її часткове або повне оновлення. В першому випадку під впливом спеціальних ферментів оновлюються невеликі ділянки поліпептидних ланцюгів або окремі амінокислотні залишки (транспептидація). В другому випадку відбувається повна заміна „зношеної молекули” білка новою. Пошкоджена молекула білка розпадається під впливом тканинних протеаз або катепсинів I, II, III і IV, що локалізуються в лізосомах. Молекула протеїду розпадається на простетичну групу і простий білок, які піддаються звичайним для цих речовин перетворенням.
Швидкість оновлення білків різних органів і тканин неоднакова. Білки організму людини в цілому оновлюються протягом 135 – 155 діб. Білки печінки, підшлункової залози, стінки кишок і плазми крові оновлюються протягом 10 діб, м'язів – 30, колаген – 300 діб. Синтез молекули білка в клітині протікає швидко – протягом 2 – 5 с. За 1 с в тілі людини руйнується близько 3 млн. еритроцитів, а один еритроцит містить близько 280 млн. молекул гемоглобіну. В організмі дорослої людини щодоби синтезується 90 – 100 г білка (1,3 г на 1 кг маси). Ступінь оновлення зменшується при старінні, хворобах і т.д.
Біосинтез пептидів. Частина ендо- і екзогенних амінокислот йде на синтез пептидів.
Глутатіон. Це трипептид, утворений із залишків глутамінової кислоти, цистеїна і гліцина.
Біосинтез протікає в дві стадії. Так, спочатку під впливом ферменту g-глутамілцистеїнсинтетази утворюється дипептид, потім за участю трипептидсинтетази – трипептид глутатіон:
L-Глутамінова кислота + L-цисцеїн + АТФ D g-Глутамілцистеїн + АДФ + Н3РО4;
g-Глутамілцистеїн + Гліцин + АТФ ® Глутатіон + АДФ + Н3РО4.
Глутатіон (G–SH) міститься у всіх тканинах і клітинах. Йому належить ведуча роль в окисно-відновних реакціях. Він є складовою частиною багатьох ферментів, захищає SH-групи білків від окислення (див. ферменти). Входить до складу тканинних катепсинів.
Карнозин і ансерин. Дипептиди м'язової тканини. Карнозин утворюється з гістидина і b-аланіна, ансерин – з 1-метил-гістидина і b-аланіна:
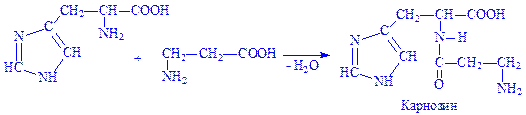
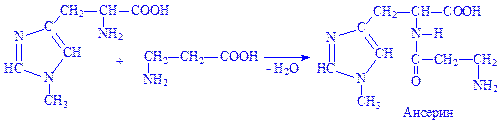
Пептиди синтезуються під впливом специфічних ферментів, за участю АТФ і Mg2+ іонів. Реакції протікають в дві стадії, наприклад синтез карнозина:
b-Аланін + АТФ + Фермент D Фермент-b-аланінаденілат + Пірофосфат;
Фермент-b-аланінаденілат + L-Гістидин D АМФ + Фермент + Карнозин
Біосинтез окремих амінокислот. Замінні амінокислоти синтезуються в тканинах організму. Незамінні амінокислоти поступають в організм у складі їжі і кормів. Умовно замінні амінокислоти синтезуються в тканинах обмежено (аргінін і гістидин) або за наявності попередників (тирозин і цистеїн). Деяка кількість амінокислот синтезується симбіотичною мікрофлорою в харчовому каналі. Синтез і обмін амінокислот – багатоступінчатий процес, що складається з ряду стадій і каталізується багатьма ферментними системами.
Матеріалом для синтезу амінокислот найчастіше є a-кето- і a-оксикислоти, які утворюються в тканинах при проміжному обміні вуглеводів, ліпідів і інших сполук. Джерелом азоту є аміак і амонійні солі, водню – НАД×H2 або НАДФ×H2.
Якщо джерелом амінокислоти є кетокислота, то вона може піддаватися відновному амінуванню, яке протікає в дві стадії: спочатку утворюється імінокислота, потім – амінокислота:
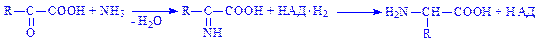
Так утворюється аланін з піровиноградної кислоти.
Щавелевооцтова кислота – джерело утворення аспарагінової і глутамінової кислот:
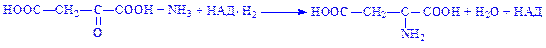
Частина глутамінової кислоти може синтезуватися з a-кетоглутарової кислоти під дією ферменту L-глутаматдегідрогенази:
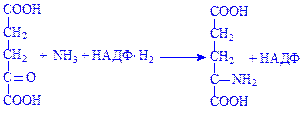
Глутамінова кислота використовується тканинами як донор аміногрупи.
Окремі амінокислоти можуть утворюватися з інших амінокислот трансамінуванням під впливом ферментів амінофераз або трансаміназ:
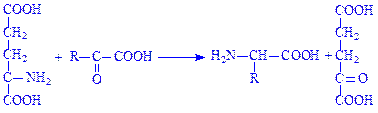
Так утворюється гліцин з серина або треоніна; аланін – з глутамінової і аспарагінової кислот, триптофана або цистеїна; тирозин з фенілаланіна; цистеїн і цистин – з серина або метіоніна; глутамінова кислота – з проліна або аргініна та ін.
Молекулярні механізми специфічності біосинтезу білків. Генетичний код. Як зазначалось вище, синтез білка відбувається на рибосомах ферментативним шляхом відповідно до інформації, закладеної в структурі ДНК. Саме ДНК є матрицею, штампом, у якому запрограмовано відповідну первинну структуру білка, що синтезується. Відомо, що ДНК знаходиться в ядрі клітини, а синтез білка відбувається на рибосомах, тому природно виникає питання, яким шляхом ДНК впливає на специфічність синтезу білка, як саме структура ДНК визначає порядок розташування залишків амінокислот у полінуклеотидному ланцюгу.
Встановлено, що зв'язуючою ланкою між ДНК ядра і рибосомами – місцем біосинтезу білка – є іРНК. Вона передає інформацію від ДНК до рибосом. Цей процес полягає в тому, що на ланцюгу ДНК ядра як на матриці проходить біосинтез іРНК. При цьому іРНК точно копіює послідовність розташування нуклеотидів одного з полінуклеотидних ланцюгів ДНК з тією лише відмінністю, що на місці тимінового нуклеотиду розташовується уридиловий нуклеотид.
Вважають, що на поверхні ДНК, згідно з законом комплементарності, може синтезуватися кілька молекул іРНК. Ділянка, на якій синтезується одна молекула PHK, називається структурним геном, або цистроном.
іРНК, отримавши інформацію від ДНК, у процесі біосинтезу переходить з ядра до рибосом, де бере безпосередню участь у формуванні первинної структури відповідного білка.
Виникає питання, як саме послідовність чотирьох різних нуклеотидів, що становлять структуру нуклеїнових кислот, визначає послідовність розташування 20 залишків амінокислот у білку. Як було розшифровано код білкового синтезу? На перший погляд можливість передачі невеликою кількістю умовних знаків інформації значної кількості можливих комбінацій залишків амінокислот у білкових молекулах може здатись сумнівною. Однак незаперечним є той факт, що всього 32 букви алфавіту передають всю красу і багатство нашої мови, і ці 32 букви можуть бути закодовані азбукою Морзе – комбінацією лише двох знаків – крапки і тире.
Використовуючи чотирьохбуквений алфавіт азотистих основ, можна скласти 4 однобуквених слова – сиглетний код, 16 двобуквених (4 ´ 4 = 16) – дуплетний код або 64 трибуквених слова (4 ´ 4 ´ 4 = 64) – триплетний код. Для 20 амінокислот шістнадцяти двобуквених слів мало, а 64 трибуквених достатньо. У 1954 р. було запропоновано кодове слово з трьох азотистих основ – триплетний код. Однак необхідно було вивчити, які нуклеотиди і в якому порядку у молекулі ДНК (іРНК) відповідають різним амінокислотам. Ці дослідження провели M. Шренберг і Г. Корана, за що були удостоєні Нобелівської премії.
Вчені добули синтетичний PHK-полімер – поліуридилову кислоту. При добавлянні її в систему, де проходив синтез білка, відбувався синтез поліфенілаланіну, а інші 19 амінокислот не включалися в полінуклеотидний ланцюг. Отже, було доведено, що три уридилових залишки (УУУ) кодують включення в поліпептидний ланцюг молекули білка амінокислоти фенілаланіну. Згодом було вияснено склад кодонів для інших амінокислот. Так, кодон ГГГ у структурі нуклеїнової кислоти забезпечує включення в поліпептидний ланцюг глутамінової кислоти, AAA – лізину, ЦЦЦ – проліну тощо (табл. 1).
| Перший нуклео-тид | Другий нуклеотид | Третій нуклео-тид | ||||||||||||
| У | Ц | А | Г | |||||||||||
| У | УУУ УУЦ | 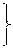
| Фен | УЦУ УЦЦ УЦА УЦГ | 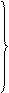
| Сер | УАУ УАЦ | 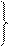
| Тир | УГУ УГЦ |
| Цис | У Ц А Г | |
| УУА УУГ | 
| Лей | УАА* УАГ* | 
| Терм | УГА* УГГ | Терм Три | |||||||
| Ц | ЦУУ ЦУЦ ЦУА ЦУГ | 
| Лей | ЦЦУ ЦЦЦ ЦЦА ЦЦГ | 
| Про | ЦАУ ЦАЦ |
| Гіс | ЦГУ ЦГЦ ЦГА ЦГГ | 
| Арг | У Ц А Г | |
| ЦАА ЦАГ |
| Глн | ||||||||||||
| А | АУУ АУЦ АУА | 
| Ілей | АЦУ АЦЦ АЦА АЦГ | 
| Тре | ААУ ААЦ |
| Асн | АГУ АГЦ |
| Сер | У Ц А Г | |
| ААА ААГ | 
| Ліз | АГА АГГ |
| Арг | |||||||||
| АУГ | – | Мет | ||||||||||||
| Г | ГУУ ГУЦ ГУА ГУГ | 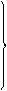
| Вал | ГЦУ ГЦЦ ГЦА ГЦГ | 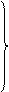
| Ала | ГАУ ГАЦ | 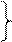
| Асп | ГГУ ГГЦ ГГА ГГГ | 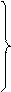
| Глі | У Ц А Г | |
| ГАА ГАГ | 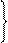
| Глу |
Таблиця 1. Генетичний код
Наведені в таблиці дані свідчать, що амінокислоти мають по кілька кодонів. Слід наголосити, що з 64 кодонів 61 визначає послідовність розміщення залишків амінокислот у поліпептидному ланцюгу, при цьому кодони ГУГ (валіновий), АУГ (метіоніновий) є також ініціюючими, стартовими кодонами. Три інших кодони – УАГ, УАА, УГА – називають беззмістовними, вони виконують функцію сигналів про закінчення синтезу поліпептидного ланцюга. В таблиці вони позначені зірками.
Аналізуючи таблицю генетичного коду, можна помітити, що перші два нуклеотиди в кодоні важливіші, ніж третій. Деякі дослідники вважають, що заміна третього нуклеотиду в значній частині кодонів не впливає на їх здатність кодувати відповідну амінокислоту. Наприклад, кодування аланіну здійснюється кодонами ГЦУ, ГЦЦ, ГЦА, ГЦГ, які відрізняються між собою третім нуклеотидом.
У зв'язку з високою специфічністю перших двох нуклеотидів у кодоні, Ф. Крік у 1965 р. запропонував теорію неоднозначної відповідності, що в точному перекладі означає коливання. З неї випливає, що головне значення при „впізнаванні” кодона антикодоном на рибосомі мають перші два нуклеотиди. Вони повністю підпорядковані комплементарному принципу взаємодії азотистих основ. Третій нуклеотид кодона може вступати у взаємодію більш ніж з одним типом нуклеотидів антикодона. Це сприяє підвищенню стійкості генетичної інформації при пошкодженні ДНК.
Особливості генетичного коду. Насамперед, слід зазначити, що код, очевидно, є універсальним. Він однаковий для всіх організмів – бактерій, рослин і тварин. Правда, одні організми віддають перевагу одним триплетам, другі – іншим. Універсальність коду свідчить про те, що він з'явився на ранніх етапах розвитку живих систем і, очевидно, мало змінюється в ході еволюції.
Іншою характерною особливістю коду є його виродженість, оскільки два і більше триплетів можуть кодувати одну амінокислоту. Хоча код і вироджений, однак він не є неоднозначним. Жоден кодон, крім ініціюючих ГУГ і АГУ, не кодує більше ніж одну амінокислоту.
У більшості випадків виродженість коду зумовлює наявність декількох тРНК для однієї амінокислоти, які відрізняються між собою антикодонами.
Особливістю коду є те, що він не перекривається, тобто нуклеотиди, які входять до складу триплету, не беруть участі в формуванні наступних триплетів. Після зчитування інформації з одного триплету механізм зчитування переміщується відразу на всі три нуклеотиди.
Характерною особливістю коду є його неперервність. Кожний триплет, який кодує відповідну амінокислоту, перебуває поряд з іншим триплетом без проміжних розділяючих ділянок (факторів).
Досить важливою особливістю коду є його однонапрямленість. Кодони під час синтезу білка транслюються в одному напрямку, починаючи з першої нуклеотидної основи. Перша азотиста основа знаходиться біля 5'-нуклеотидного кінця, а остання – біля 3'-кінця, тобто зчитування інформації проходить у напрямку 5' ® 3'.
Слід зазначити також, що протягом останніх років з'явились дані, які свідчать про окремі відхилення від загальноприйнятих особливостей генетичного коду. Так, Б. Берел із співробітниками, вивчаючи нуклеотидну послідовність ДНК мітохондрій людини, встановили, що в цілому в них генетичний код подібний до встановленого раніше. Разом з тим виявлено, що чотири кодони змінили свій зміст. Кодон УГА відповідає триптофану, кодон АУА – метіоніну, а кодони – АГА і АГГ стали термінуючими. В мітохондріях дріжджів всі чотири лейцинових кодони, які починаються з ЦУ, перейшли до треоніну. Отже, у лейцину залишилось лише два кодони, а у треоніну їх стало шість. В одній клітині, зокрема організму людини, виявлено існування двох різних кодів. Ці дані, очевидно, є доказом того, що код зазнав певних, незначних еволюційних змін, і універсальність коду має свої нюанси.
Потребує певних корективів і особливість коду, що він не перекривається. В дослідах з вірусами і деякими бактеріями встановлено, що одна і та сама ділянка ДНК може кодувати кілька різних білків за принципом зміщення рамки зчитування. Так, якщо взяти будь-який код з певною послідовністю нуклеотидів, наприклад ТАГАТГЦГЦА, то при зчитуванні його з першої букви дістанемо триплети ТАГ, АТГ, ЦГЦ, а при зчитуванні з другої букви – зовсім інші триплети: АГА, ТГЦ, ГЦА і т. д. Разом з цим встановлено, що для окремих білків частина послідовностей нуклеотидів спільна. Наприклад, якщо для одного білка дана послідовність нуклеотидів закінчується, то для іншого вона може продовжуватися. Одержані дані свідчать про те, що транскрипція генетичного коду підпорядкована певним контрольним механізмам, які забезпечують вибір певного правильного коду.
Встановлено ряд нових фактів, які підтверджують, що в генах еукаріот є некодуючі ділянки. Іншими словами, гени вищих організмів не неперервні, а побудовані з окремих частин, розділених іншими послідовностями нуклеотидів. Проміжки між цими частинами бувають різними – від 10 до 20 000 пар нуклеотидів.
Генна інженерія і біосинтез білка. Генна інженерія відкриває нову еру в біології. Це пояснюється насамперед тим, що з'явилися нові можливості для проникнення в глибину біологічних явищ з метою повнішої характеристики форм існування живої матерії, ефективнішого вивчення структури і функцій генів на молекулярному рівні для розуміння тонких механізмів роботи генетичного апарата. Разом з цим генна інженерія забезпечує моделювання на молекулярному рівні мутацій і рекомбінацій – основних генетичних процесів, факторів еволюції.
Успішне проведення досліджень у галузі генної інженерії має крім теоретичного важливе практичне значення. Генна інженерія створює основи для пізнання способів та шляхів „конструювання” нових або направленої зміни існуючих організмів, сприяє позитивному розв'язуванню проблеми направленого біосинтезу важливих для організму сполук, зокрема білків. Суть цього процесу полягає в тому, що за допомогою певних методів (синтез на іРНК за участю ревертази, звичайний хімічний синтез) добувають ген, який визначає синтез відповідного білка. Далі такий ген вбудовується у відповідну систему, яка дає йому можливість швидко розмножуватися. Такою системою, яку часто називають вектором, служать плазміди – автономні структури, що несуть свою інформацію у вигляді невеликої циклічної молекули ДНК. Вони виявлені в клітинах бактерій разом з бактеріальною хромосомою. Крім плазмід можна використовувати також інші системи – віруси, фаги і т.д.
Плазміди, що містять вбудований ген (рекомбінантні ДНК), вводять у клітини бактерій, які завдяки цьому набувають здатності синтезувати нові для них речовини. Це основний принцип генної інженерії, мета якої – добування білків людини, тварин і рослин, які особливо необхідні в практичній діяльності, наприклад у медицині. До таких важливих білків належить, наприклад, гормон інсулін. Нестача інсуліну в організмі людини призводить до тяжкого і досить поширеного захворювання – діабету. Для його лікування використовують інсулін тваринного походження, добутий з підшлункової залози великої рогатої худоби або свиней.
Однак останнім часом з'ясовано, що для частини людей він не ефективний або ж викликає сильну алергію. Це пояснюється тим, що інсулін людського і тваринного організмів характеризується певною структурною відмінністю. Отже, виникла потреба у виробництві інсуліну людини. Добувати його синтетично економічно невигідно.
Генна інженерія дала рентабельніший шлях синтезу інсуліну. Добуто штами бактерій з рекомбінантними ДНК, які виробляють проінсулін людини, з якого далі легко добути інсулін.
Другим важливим здобутком генної інженерії є інтерферон – універсальний противірусний препарат. Основою його є білок, який виробляється клітинами організму для боротьби з вірусною інфекцією. Вивчення властивостей інтерферону показало, що він виявляє високу видову специфічність. Тому для лікування людей можна використовувати інтерферон лише людини.
Інтерферон добувають в основному із лейкоцитів донорської крові. Однак такий шлях добування інтерферону досить дорогий і, крім того, не здатний повністю забезпечити потребу в ньому. Тому інтенсивно ведуться роботи по розробці нових методів добування інтерферону, зокрема шляхом генної інженерії. В цьому плані програма добування штаму – продуцента інтерферону – успішно завершена, що дало змогу виробляти препарат у достатніх кількостях. Крім того, методом генної інженерії добуто ще один важливий білок – соматотропін. Це гормон росту людини, до складу якого входить 191 залишок амінокислот. Соматотропін застосовують для лікування карликовості, при переломах кісток, опіках та при інших захворюваннях.
Отже, генна інженерія відкриває нову технологію (біотехнологію) синтезу важливих для організмів людини і тварин речовин, зокрема білків. Це дає можливість значно знизити вартість і розширити асортимент виробництва гормонів, ферментів та інших білкових препаратів, які використовуються в медицині та сільському господарстві.
Регуляція синтезу білка. Необхідною умовою існування живих систем, забезпечення їх життєдіяльності є наявність узгодженої системи регулювання найважливіших процесів, які протікають в них. Це, насамперед, стосується синтезу білка, оскільки оптимальне співвідношення між кількістю і якістю різних білків відіграє важливу роль у забезпеченні ряду життєво важливих процесів як для одноклітинних, так і для багатоклітинних організмів. За участю білків-ферментів здійснюється регуляція обміну речовин, адаптація організмів до зміни умов зовнішнього і внутрішнього середовищ, процеси онтогенезу та диференціації клітин, органів і систем. Тому досить важливим є здійснення контролю за синтезом саме тих білків-ферментів, які необхідні клітині за даних умов залежно від функцій, які вона повинна виконувати. В зв'язку з цим у процесі тривалого еволюційного розвитку живих організмів, незалежно від ступеня їх організації, сформувався досить складний, узгоджений механізм регуляції синтезу білка, який забезпечує сталі якісний та кількісний склад білків, необхідний для виконання певних фізіологічних функцій.
З'ясування суті механізму регуляції синтезу білка є досить складною проблемою, яка ще не повністю розв'язана. Над вирішенням цієї проблеми протягом кількох десятиріч працювала велика група різних дослідників-генетиків, біохіміків, молекулярних біологів. Логічним розвитком даних досліджень стала концепція регуляції синтезу білка, розроблена в 1961 р. лауреатами Нобелівської премії французькими вченими Ф. Жакобо і Ж. Моно. Ґрунтується вона на спостереженнях за індукцією і репресією утворення ферментів у клітинах бактерій E. соlі.
Дослідженнями виявлено збільшення концентрації деяких ферментів бактерій при добавлянні в середовище субстратів, на які вони діють, і зниження концентрації ферментів при наявності кінцевих продуктів реакцій, що каталізуються даними ферментами. Отже, між цими процесами встановлено тісний взаємозв'язок.
Концепція Ф. Жакобо і Ж. Моно, сформульована в дослідах на прокаріотах, дістала широке визнання і вважається загальноприйнятою. Згідно з цією концепцією, регуляція синтезу білка здійснюється на рівні ДНК, молекула якої складається з певних функціональних ділянок – генів, згрупованих залежно від функцій, які вони виконують. Одна група таких функціональних ділянок названа структурними генами, або цистронами, вони містять інформацію про синтез певних поліпептидних ланцюгів білка, на них відбувається синтез молекул іРНК, яка далі надходить до рибосом, де виконує роль матриці в процесі синтезу білка. Початок синтезу іРНК – зчитування генетичної інформації – розпочинається з функціональної ділянки ДНК, яка має назву промотора і є точкою ініціації її синтезу.
Іншу групу функціональних ділянок ДНК становлять регуляторні гени, які регулюють активність структурних генів шляхом їх „включення” та „виключення”. Регуляторні гени представлені геном-оператором, що знаходиться безпосередньо біля групи структурних генів, і геном-регулятором, що знаходиться від них на деякій відстані.
Ген-оператор є ніби пусковим механізмом, який, залежно від умов, дозволяє або гальмує синтез іРНК на структурних генах ДНК. Вважають, що ген-оператор локалізований на крайньому відрізку цистронів і є, очевидно, вихідним пунктом дії ДНК-залежної РНК-полімерази.
Гени-оператори разом з групами структурних генів утворюють групи узгоджено діючих блоків – оперонів, кожен з яких відповідає за взаємозв'язаний синтез ряду специфічних білків, тобто оперон є одиницею транскрипції. Якщо ген-оператор не діє, то весь оперон стає бездіяльним – гальмується синтез іРНК, а отже, і білків, ферментів. Діяльність гена-оператора регулюється геном-регулятором. Оскільки ген-регулятор і структурні гени оперона розташовані на різних ділянках ДНК, то зв'язок між ними забезпечують речовини-посередники – білки-репресори, що утворюються в рибосомах на матриці специфічної мРHK, яка синтезується на гені-регуляторі.
Свою назву білок-репресор дістав завдяки тому, що його дія на ген-оператор гальмує (репресує) діяльність останнього, внаслідок чого припиняється функціонування всієї групи структурних генів. Білки-репресори виявляють спорідненість до гена-оператора і здатність до зворотного зв'язування з ним. Крім того, білок-репресор здатний до специфічного зв'язування з певними низькомолекулярними речовинами, так званими індукторами, або ефекторами.
За звичайних фізіологічних умов репресори можуть перебувати в активному або пасивному стані. Перехід з одного стану в інший регулюється продуктами внутрішньоклітинного обміну або речовинами, що надходять із зовнішнього середовища.
Залежно від стану білка-репресора розрізняють індуцибельну і репресибельну системи генної регуляції синтезу білка.
При індуцибельній системі регуляції білок-репресор, що утворюється на гені-регуляторі, перебуває в активному стані. Його вплив на ген-оператор призводить до блокування діяльності оперона і припинення синтезу іРНК і певних білків-ферментів. Синтез їх може відновлюватися лише тоді, коли в клітині з'являються продукти, для утилізації яких необхідні дані ферменти. Білок-репресор, сполучаючись з цими продуктами (індукторами), втрачає здатність контролювати ген-оператор, внаслідок чого відновлюється синтез іРНК.
Вважають, що індуктор, сполучаючись з білком-репресором, зумовлює зміну його третинної структури, внаслідок чого останній втрачає здатність до зв'язування з субстратом – геном-регулятором. Разом з тим оператор, який виходить з-під контролю гена-регулятора, набуває активності і приводить у дію блок структурних генів, на яких здійснюється синтез іРНК, необхідних для утворення певних ферментів. Оскільки дані ферменти спрямовані на утилізацію Індуктора, білок-репресор буде пасивним доти, поки під впливом ферментів не відбудеться повне розщеплення індуктора. Після цього білок-репресор вивільнюється, переходить в активний стан і блокує оперон, внаслідок чого синтез іРНК, що кодує первинну структуру даних ферментів, припиняється.
Крім індукції генів у клітинах відбувається їх репресія. Особливо часто цей процес спостерігається при процесах синтезу, коли концентрація багатьох ферментів значно знижується при збільшенні концентрації кінцевих продуктів реакцій, що каталізуються даними ферментами. При репресибельній системі регуляції синтезу білка білок-репресор, що синтезується на рибосомах клітини, перебуває в пасивному стані і не може пригнічувати діяльність гена-оператора, а отже, і не контролює діяльність оперона, на якому проходить синтез іРНК. Активація білка-репресора, блокування оперона і припинення синтезу іРНК відбуваються під впливом корепресора, який утворює комплекс з опероном. Корепресорами є кінцеві продукти реакції синтезу, або один з цих продуктів. Є дані, що корепресором у процесі синтезу ферментів обміну амінокислот може бути не вільна амінокислота, як кінцевий продукт реакції біосинтезу, а її комплекс з тРНК (аміноацил-тРНК).
Блокування гена-оператора припиняється тоді, коли вичерпується весь корепресор. Внаслідок цього за відсутності активатора (корепресора) білок-репресор переходить у свій звичайний пасивний стан, вивільнює ген-оператор і групу структурних генів, на яких відновлюється синтез білків-ферментів.
Як зазначалось раніше, концепція Ф. Жакобо і Ж. Моно відносно регуляції синтезу білка стосується нижчих організмів – прокаріот. Механізм регуляції цих процесів у клітинах високоорганізованих організмів – еукаріот – значно складніший. Певний внесок, у розв'язання цієї проблеми зробив Г.П. Георгієв. Згідно з його гіпотезою, в клітинах вищих організмів гени також згруповані в оперони, однак їхній склад не такий простий, як у мікроорганізмів. Основна відмінність полягає в тому, що оперон має ряд генів-операторів і кожен з них реагує з іншим білком-репресором. Блокування будь-якого гена-оператора перешкоджає просуванню ферменту ДНК-залежної PHK-полімерази. Як результат цього синтез іРНК на структурних генах сповільнюється або припиняється зовсім.
Із-за наявності кількох генів-операторів на активність оперона впливає кілька різних факторів. Наприклад, один ген-оператор може блокуватися з підвищенням концентрації певного гормона, інший – з нагромадженням у клітині певного продукту обміну і т.д. Крім цього, один і той самий ген-оператор входить до складу різних оперонів. Тому ці оперони реагують на один і той самий фактор. Це свідчить про те, що будь-який один фактор може регулювати відразу кілька оперонів, а активність одного оперона може залежати від кількох різних факторів. Це створює досить точну і разом з тим гнучку систему саморегуляції.
Слід зазначити, що ряд питань, які стосуються механізмів, що зумовлюють специфічність біосинтезу білків, а також регуляції синтезу білків, вивчено ще недостатньо.
Перетворення амінокислот. В основі перетворення амінокислот лежать три основні види реакцій: реакції за аміногрупою, за карбоксильною групою і за радикалом амінокислоти. Перші два види реакцій однотипні для всіх L-амінокислот. Реакції амінокислот за радикалом досить різноманітні ї специфічні майже для кожної амінокислоти.
Перетворення амінокислоти за аміногрупою. До таких реакцій належать два основні види перетворення амінокислот: дезамінування і переамінування.
Дезамінування – процес відщеплення аміногрупи від амінокислоти з утворенням молекули аміаку. Відомо чотири види дезамінування: окислювальне, відновне, гідролітичне і внутрішньомолекулярне. Під час окислювального дезамінування амінокислоти перетворюються на кетокислоти. Під час відновного дезамінування амінокислоти перетворюються на насичені жирні кислоти. Гідролітичне дезамінування пов'язане з перетворенням амінокислот на оксикислоти. Внутрішньомолекулярне дезамінування характеризується перетворенням амінокислот на ненасичені жирні кислоти (див. розклад білків).
Зазначені вище реакції каталізують специфічні ферменти. Найбільш поширеним в організмах людини і тварин є окислювальне дезамінування. Розглянемо його механізм на прикладі глутамінової кислоти. Спочатку амінокислота при каталітичній дії глутаматдегідрогенази, коферментом якої є НАД+ або НАДФ+ (для дегідрогеназ інших амінокислот коферментом є ФМН), окислюється до іміноглутарової кислоти:
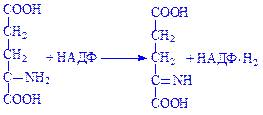
Далі іміноглутарова кислота спонтанно, без участі ферменту гідролізує з утворенням a-кетоглутарової кислоти й аміаку:
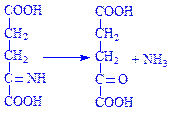
Слід зазначити, що обидві реакції оборотні. Це свідчить про те, що з a-кетоглутарової кислоти може утворюватись глутамінова кислота. Іншим видом перетворення амінокислот за аміногрупою є переамінування (трансамінування) амінокислот. Цей процес перетворення амінокислот дослідили радянські біохіміки O.Є. Браунштейн і M.Г. Кріцман у 1937 р. Вони експериментально встановили, що під час взаємодії глутамінової і піровиноградної кислот можуть утворюватись a-кетоглутарова кислота та аланін без проміжного утворення аміаку. Ці дані наштовхнули вчених на думку, що аміногрупа з амінокислоти за допомогою ферментативних систем може переноситись на кетокислоти. При цьому амінокислота перетворюється на кетокислоту, а кетокислота – в амінокислоту:
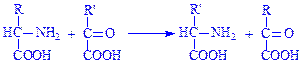
Дослідження механізму переамінування показало, що в цьому процесі беруть участь ферменти амінотрансферази, коферментом яких є фосфорний ефір вітаміну B3 (піридоксальфосфат). Під час реакції піридоксальфосфат приєднує аміногрупу амінокислоти і перетворюється на фосфопіридоксамін, після чого віддає аміногрупу кетокислоті і знову перетворюється на піридоксальфосфат. Розглянемо механізм процесу переамінування на прикладі взаємодії глутамінової і піровиноградної кислот. Для зручності піридоксальфермент позначимо:

На першій стадії відбувається взаємодія глутамінової кислоти з піридоксальферментом, в результаті чого утворюється фермент-субстратний комплекс. Далі відбувається таутомерне перегрупування комплексу, пов'язане з переміщенням атома водню від залишку глутамінової кислоти до піридоксалю, внаслідок чого подвійний зв'язок також переміщується і виникає між атомами азоту і вуглецю глутамінової кислоти:
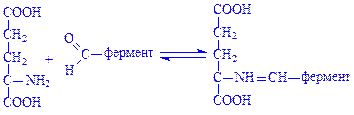

Після таутомерного перегрупування відбувається гідролітичне розщеплення фермент-субстратного комплексу з утворенням a-кетоглутарової кислоти і піридоксамін-ферменту:
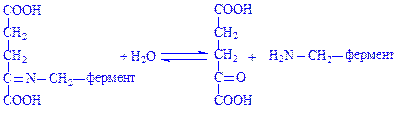
Піридоксамін-фермент, що утворився, взаємодіє з піровиноградною кислотою з утворенням іншого фермент-субстратного комплексу:
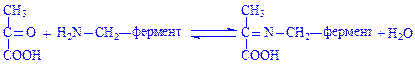
У цьому комплексі знову відбувається перегрупування:
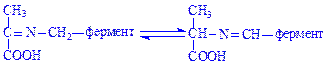
Далі відбувається гідроліз комплексу з утворенням амінокислоти (аланіну) і виділенням піридоксаль-ферменту:
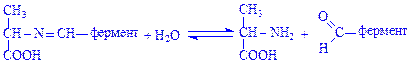
Отже, у процесі переамінування амінокислота (глутамінова) перетворилась на кетокислоту (a-кетоглутарову), а кетокислота (піровиноградна) – на амінокислоту (аланін). Переамінування амінокислот відбувається у різних органах і тканинах. Воно відіграє важливу роль у процесах синтезу необхідних для організму амінокислот. Кетокислоти, що утворюються внаслідок переамінування, можуть використовуватись для синтезу ліпідів, вуглеводів та інших речовин.
Слід зазначити, що між переамінуванням і окислювальним дезамінуванням існує певний зв'язок. Він полягає в існуванні непрямого дезамінування, тобто в перенесенні аміногрупи a-амінокислоти на a - кетоглутарову кислоту шляхом трансамінування. При цьому a-амінокислота перетворюється на кетокислоту, а глутамінова кислота піддається дезамінуванню з утворенням a-кетоглутарової кислоти й аміаку:
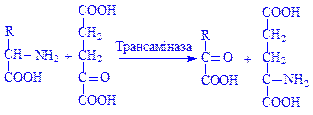

Зазначені реакції свідчать про те, що дезамінування вихідної амінокислоти відбувається не безпосередньо, а через трансамінування за участю a-кетоглутарової кислоти як акцептора аміногрупи.
Перетворення амінокислот за карбоксильною групою. До таких реакцій належать реакції декарбоксилування амінокислот з утворенням аміноациладенілатів.
Процес декарбоксилування амінокислот досить поширений як у тваринних, так і в рослинних організмах. Він пов'язаний з відщепленням від молекули амінокислоти CO2 за рахунок карбоксильної групи, в результаті чого утворюються відповідні аміни.
Реакції декарбоксилування каталізують ферменти декарбоксилази. У вигляді коферменту в їх складі виступає піридоксальфосфат. Винятком є декарбоксилаза гістидину.
Аміни, що утворюються в процесі декарбоксилування, мають високу біологічну активність, тому їх називають також біогенними амінами. Наприклад, під час декарбоксилування глутамінової кислоти утворюється g-аміномасляна кислота:
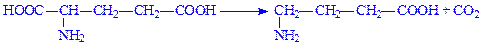
Ця амінокислота відіграє важливу роль у функціональній діяльності центральної нервової системи.
Іншим прикладом є декарбоксилування гістидину з утворенням гістаміну:
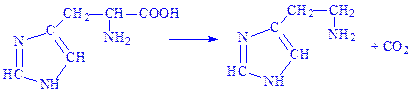
Гістамін здатний розширювати судини, знижувати кров'яний тиск, а також активізувати діяльність залоз внутрішньої секреції.
Слід зазначити, що аміни виявляють фізіологічну дію при досить малих концентраціях.
Нагромадження їх в організмі може викликати порушення ряду біохімічних процесів. Щоб запобігти цьому, в тканинах є активна амінооксидаза, яка каталізує процеси окислення амінів до альдегідів і кислот, частина яких виводиться з організму з сечею, а інша частина піддається перетворенням.
Процеси декарбоксилування більш характерні для мікроорганізмів і протікають дещо обмежено в організмах людини і тварин.
Іншою характерною реакцією амінокислот за карбоксильною групою є утворення аміноациладенілатів.
Реакція відбувається за участи специфічного ферменту і АТФ:
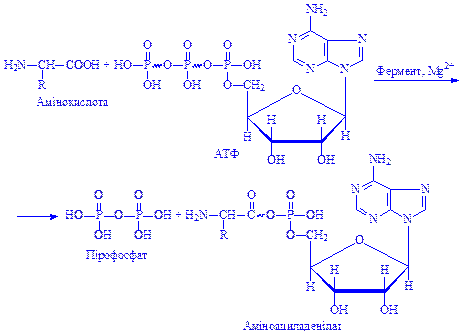
Утворення аміноациладенілатів має важливе значення для активації амінокислот, які включаються в процес біосинтезу білка.
Перетворення амінокислот за радикалами. Для амінокислот, крім реакцій за аміно- та карбоксильною групами, властиві хімічні перетворення, пов'язані з реакціями за радикалом.
Характерною особливістю радикалів амінокислот є те, що вони містять різні функціональні групи, рухомі атоми водню тощо. Цим зумовлена активна участь їх в багатьох хімічних реакціях, частина з яких приводить до утворення ряду замінних амінокислот в організмі.
Для прикладу розглянемо реакції окислення окремих амінокислот. Так, під час окислення радикала амінокислоти фенілаланіну утворюється амінокислота тирозин:
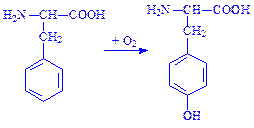
Реакцію каталізує фермент фенілаланін-гідроксилаза.
Активну участь в окисно-відновних процесах беруть радикали сірковмісних амінокислот – цистеїну і цистину. Наприклад, під час окислення двох молекул цистеїну утворюється молекула амінокислоти цистину:
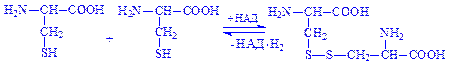
Реакція відбувається за участю ферменту цистеїнредуктази, коферментом якої є НАД+.
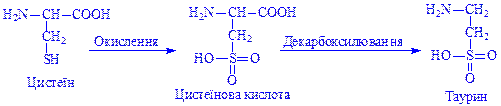
При повному окисленні сульфгідрильної групи цистеїну утворюється цистеїнова кислота, яка під час декарбоксилування перетворюється на таурин:
Важливе значення для організму мають реакції трансметилування, які здійснюються за участю АТФ. У цих процесах універсальним донором метальної групи є амінокислота метіонін. Спочатку метіонін взаємодіє з АТФ, внаслідок чого утворюється S-аденозилметіонін – активна форма метіоніну:
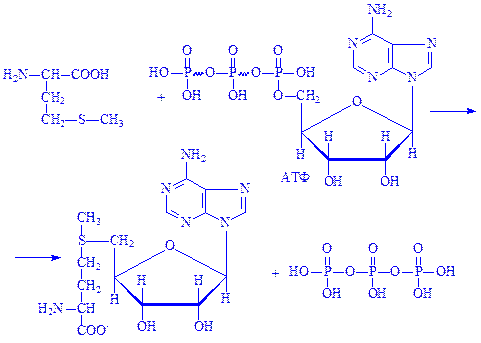
Цю реакцію каталізує фермент S-аденозилметіонінсинтетаза. Далі метильна група за участю S-метиладенозилметіоніну переноситься на сполуку, яка піддається метилюванню, наприклад на диметиламіноетанол. При цьому утворюється холін і S-аденозилгомоцистеїн. Реакція каталізується метилтрансферазою:
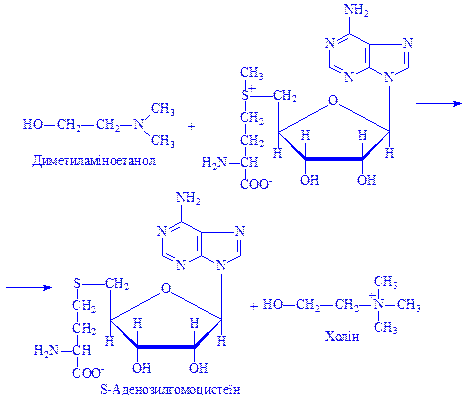
Іншим прикладом перетворення амінокислот за радикалом може бути гідролітичне розщеплення аргініну за участю ферменту аргінази. При цьому утворюється амінокислота орнітин і сечовина:
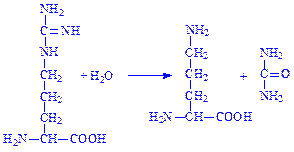
Слід підкреслити що радикалу кожної амінокислоти властиві певні хімічні перетворення.
Обмін окремих амінокислот має індивідуальні особливості.
Гліцин. Бере участь у ряді найважливіших реакцій біосинтезу. Так, з нього утворюються:
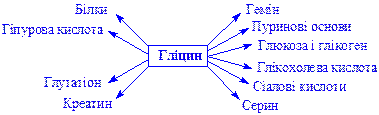
У тканинах печінки гліцин бере участь в процесах знешкодження отруйних сполук – бензойної, фенілоцтової кислот і фенолів, утворює парні сполуки, які виводяться з сечею з організму.
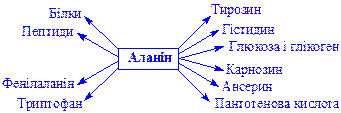
Аланін. Утворюється трансамінуванням піровиноградної кислоти (див. вище). Існує у вигляді a- і b-форм. Бере участь у біосинтезі:
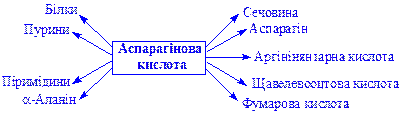
Аспарагінова кислота. Утворюється трансамінуванням щавелевооцтової кислоти (див. вище). Разом з глутаміновою кислотою забезпечує взаємозв'язок між обміном білків, вуглеводів і ліпідів. Служить донатором аміногруп в реакціях трансамінування. Основні реакції відображає схема:
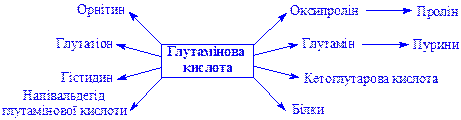
Глутамінова кислота. Міститься в тканинах у складі білків, у вільному стані і у вигляді аміда. Донатор аміногрупи в реакціях трансамінування. Основні речовини, в синтезі яких бере участь кислота:
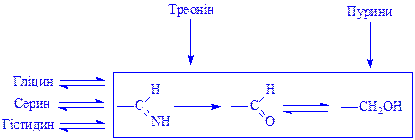
Серин і треонін. Їх обмін тісно пов'язаний з обміном гліцина. Серин у тканинах утворюється з 3-фосфогліцеринової кислоти. З серина утворюється гліцин у результаті перенесення одновуглецевого фрагмента (C1) на тетрагідрофолієву кислоту (ТГФК). Гліцин може утворюватися з треоніна. Фрагмент C1 використовується для синтезу гістидина і пуринів. З серина і треоніна утворюється піровиноградна кислота, яка за допомогою ацетил-КоА включається в ЦТК. Частину перетворень відображає схема:
Гідроксильна група серина входить до складу активного центру багатьох ферментів: трипсину, хімотрипсину, естераз, фосфорилаз, фосфатаз.
Метіонін. Є складовою частиною багатьох білків. Служить донатором метильної групи для утворення в організмі багатьох важливих сполук. Передача метильної групи в процесі переметилування відбувається під впливом відповідних метилтрансфераз через S-аденозилметіонін:
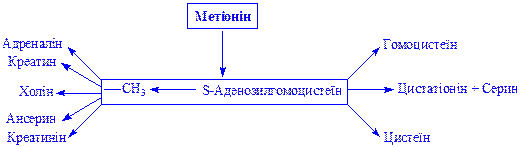
Попередником метіоніна є аспарагінова кислота, яка під впливом ряду ферментів через декілька стадій (гомосерин, О-сукциніл-гомосерин, цистеїн, цистатіонін, гомоцистеїн) перетворюється в метіонін.
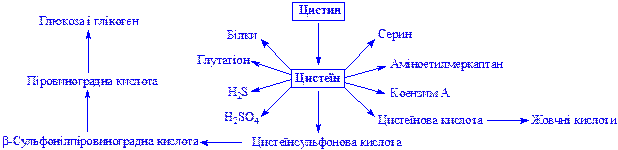
Цистеїн і цистин. Складові частини багатьох білків, пептидів, гормонів і інших сполук. SH-група цистеїну – складова частина активних центрів ряду ферментів. Участь цистеїну в обміні речовин частково відображає схема:
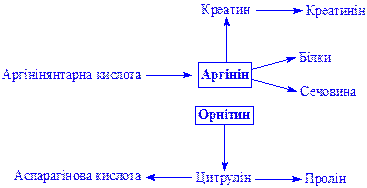
Аргінін і орнітин. Аргінін утворюється в процесі перетворення вуглекислого газу і аміаку в сечовину. Обидві амінокислоти беруть участь в утворенні ряду життєво важливих речовин:
Лізин. Найважливіша амінокислота. Бере участь у синтезі багатьох речовин:
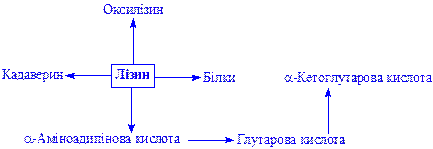
S-Аміногрупа залишку лізину бере участь у формуванні зв'язку між апо- і коферментами, особливо при утворенні біотин-ферменту. Лізину належить важлива роль в скріпленні фосфору при мінералізації кісткової тканини і інших процесах.
Фенілаланін і тирозин. У організмі ці амінокислоти використовуються при біосинтезі білків і пептидів, утворенні протеїногенних амінів, гормонів і пігментів, піддаються окисленню до кінцевих продуктів з розривом ядра та ін.:

Триптофан. ого перетворення ілюструються схемою:
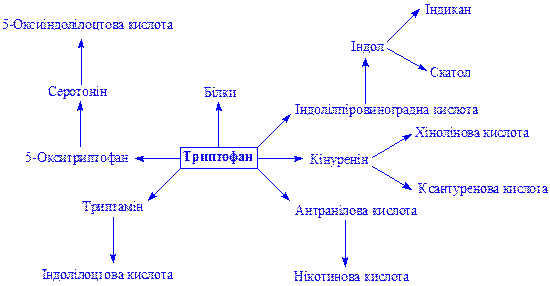
Гістидин. Відноситься до незамінних амінокислот. Бере участь в біосинтезі і обміні багатьох життєво важливих речовин:
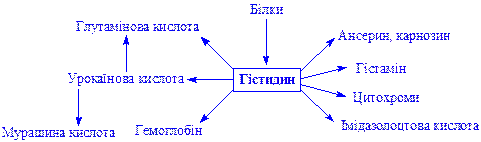
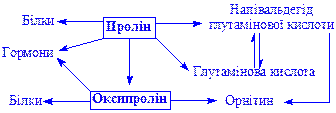
Пролін і оксипролін. Оксипролін утворюється з проліну. Процес необоротний. Обидві імінокислоти використовуються для біосинтезу білків, пептидів, гормонів і інших речовин:
Перетворення безазотистого залишку амінокислот. Частина амінокислот, які не використані в синтезі білків і їх похідних, піддається необоротним процесам розпаду до аміаку і карбонових кислот. Аміак знешкоджується у печінці в орнітиновому циклі. З декількох видів дезамінування, що протікають в органах і тканинах організму, переважає окислювальне дезамінування. Кетокислоти, що утворилися при цьому можуть використовуватися тканинами для різних потреб.
За напрямом використання безазотистого залишку амінокислоти ділять на дві групи: глюкопластичні і ліпопластичні. З глюкопластичних амінокислот (аланін, серин, цистеїн, ін.) утворюється піровиноградна кислота, яка використовується як початкова речовина для біосинтезу глюкози і глікогену.
З ліпопластичних амінокислот (лейцин, ізолейцин, аргінін, орнітин, лізин, оксилізин, ін.) після дезамінування утворюється ацетооцтова кислота – джерело біосинтезу вищих жирних кислот.
a-Кетокислоти, що утворилися при окислювальному дезамінуванні амінокислот, декарбоксилуються і одночасно окислюються в жирні кислоти. Жирні кислоти, що утворилися, можуть піддаватися b-окисленню, де кінцевим продуктом є ацетил-КоА – джерело хімічної енергії або сировина для біосинтезу багатьох речовин.
Особливості проміжного обміну складних білків. В організмі синтезуються численні складні білки, які беруть участь в різних реакціях обміну, що визначають існування живої матерії. Біосинтез складних білків протікає аналогічно біосинтезу протеїнів. При цьому формуються первинна, вторинна, третинна і четвертна структури білкової молекули, з приєднанням відповідної простетичної групи: для нуклеопротеїдів – ДНК або РНК, хромопротеїдів – гема або іншої речовини, ліпопротеїдів – ліпіду, глюкопротеїдів – вуглеводу, фосфопротеїдів – фосфорної кислоти.
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 2019; Нарушение авторских прав?; Мы поможем в написании вашей работы!