КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Мутации
|
|
|
|
Мутации (от лат. mutatio — перемена) — это изменение генов и хромосом, фенотипически проявляющиеся в изменении свойств и признаков организмов. Впервые их описал в 1901 г. голландский ученый Г. Де Фриз (1848-1935). Он же заложил основы и теории мутаций. Процесс образования мутаций во времени и пространстве называют мутагенезом. Мутации характерны для всех живых существ, включая человека. Определяя мутации и мутагенез, следует также определить и такие понятия, как дикий тип организма или гена и мутантный организм (мутант). Под диким типом понимают организмы со свойствами, находимыми в природе. Диким типом обозначают также наборы генов или отдельные гены, также являющиеся природными для организма. Образно говоря, дикий тип — это «справочный» тип, своего рода эталон, с которым сравнивают мутантные организмы и мутантные гены.
Мутантные организмы (мутанты) — это организмы, содержащие в своем геноме одну или несколько мутаций. Мутантные организмы могут отличаться от исходных (организмов дикого типа) по самым различным свойствам — морфологическим, физиологическим, биохимическим и другим. Например, у микроорганизмов мутации сопровождаются изменением формы колоний, питательных потребностей, отношения к лекарственным веществам и т. д. У высших растений мутации сопровождаются изменениями качественных и количественных признаков (рис. 115). У насекомых мутанты отличаются от исходных организмов по форме и окраске тела, крыльев, конечностей, глаз, реакции на свет, серологическим свойствам и т. д. (рис. 116). У млекопитающих мутации также ведут к изменению многих качественных и количественных признаков (рис. 117). У человека мутации приводят к различным отклонениям от нормы и сопровождаются наследственной патологией (см. гл. XIII). В целом можно сказать, что мутации являются вредными, полезными или нейтральными для организмов. Но важно помнить, что мутанты остаются организмом того же вида, что и организм дикого типа, из которого они произошли.
Мутации следует отличать от фенокопий, которые продуцируются факторами среды. Мимикрируя действие генов, они не передаются по наследству. Например, если беременных мышей экспонировать к пониженному атмосферному давлению, то некоторые из особей в пометах таких мышей будут иметь повреждения в мочеполовом тракте, не передающиеся, однако, по наследству. В случае человека фенокопиями можно считать, например, ненаследуемые повреждения скелета и слепоту, вызываемые ядами и другими вредными факторами воздействия.
Мутации возникают на всех стадиях индивидуального развития организмов и поражают гены и хромосомы как в половых клетках, причем до оплодотворения и после оплодотворения (после первого деления оплодотворенных яйцеклеток), так и в соматических, причем в любой фазе клеточного цикла. Поэтому по типу клеток, в которых возникают мутации, различают генеративные и соматические мутации (соответственно).

Генеративные мутации происходят в генитальных и половых клетках.
Если мутация (генеративная) происходит в генитальных клетках, то мутантный ген могут получить сразу несколько гамет, что увеличит потенциальную способность наследования этой мутации несколькими особями (индивидуумами) в потомстве. Если мутация произошла в гамете, то, вероятно, лишь одна особь (индивид) в потомстве получит этот ген. На частоту мутаций в половых клетках оказывает влияние возраст организма.
Соматические мутации встречаются в соматических клетках организмов. У животных и человека мутационные изменения будут сохраняться только в этих клетках. Но у растений из-за их способности к вегетативному размножению мутация может выйти за пределы соматических тканей. Например, знаменитый зимний сорт яблок «Делишес» берет начало от мутации в соматической клетке, которая в результате деления привела к образованию ветви, имевшей характеристики мутантного типа. Затем следовало вегетативное размножение, позволившее получить растения со свойствами этого сорта.
По типу наследования различают доминантные, полудоминантные, кодоминантные и рецессивные мутации. Доминантные мутации характеризуются непосредственным эффектом на организм, полудоминантные мутации заключаются в том, что гетерозиготная форма по фенотипу является промежуточной между формами АА и аа, а для кодоминантных мутаций характерно то, что у гетерозигот A1A2 проявляются признаки обоих аллелей. Рецессивные мутации не проявляются у гетерозигот.
Если доминантная мутация встречается в гаметах, ее эффекты выражаются непосредственно в потомстве. Многие мутации у человека являются доминантными. Они часты у животных и растений. Например, генеративная доминантная мутация дала начало анконской породе коротконогих овец.
Примером полудоминантной мутации может служить мутационное образование гетерозиготной формы Аа, промежуточной по фенотипу между организмами АА и аа. Это имеет место в случае биохимических признаков, когда вклад в признак обоих аллелей одинаков.
Примером кодоминантной мутации являются аллели IA и IB, детерминирующие группу крови IV.
В случае рецессивных мутаций их эффекты скрыты в диплоидах. Они проявляются лишь в гомозиготном состоянии. Примером являются рецессивные мутации, детерминирующие генные болезни человека.
Таким образом, главными факторами в детерминировании вероятности проявления мутантного аллеля в организме и популяции являются не только стадия репродуктивного цикла, но и до-минантность мутантного аллеля.
В зависимости от локализации в клетках различают генные (точечные) и хромосомные мутации (рис. 118).
Генные мутации заключаются в изменениях индивидуальных генов. Поэтому их еще называют точечными мутациями и классифицируют на односайтовые и многосайтовые. Односайтовая мутация затрагивает один сайт, многосайтовая — несколько сайтов генного локуса. Некоторые сайты являются «горячими точками», т. к. в них происходят концентрированные мутации, что связано с наличием в нуклеотидных последовательностях модифицированных оснований. Последние подвергаются частому дезаминированию, а это ведет к изменениям в последовательностях оснований.
Генные мутации классифицируют также на прямые и обратные (реверсивные) мутации, которые одинаково встречаются у организмов всех систематических групп.
Прямые мутации — это мутации, инактивирующие гены дикого типа, т. е. мутации, которые изменяют информацию, закодированную в ДНК, прямым образом, в результате чего изменение от организма исходного (дикого) типа идет прямым образом к организму мутантного типа.
Обратные мутации представляют собой реверсии к исходным (диким) типам от мутантных. Эти реверсии бывают двух типов. Одни из реверсий обусловлены повторными мутациями аналогичного сайта или локуса с восстановлением исходного фенотипа и их называют истинными обратными мутациями. Другие реверсии представляют собой мутации в каком-то другом гене, которые изменяют выражение мутантного гена в сторону исходного типа, т. е. повреждение в мутантном гене сохраняется, но он как бы восстанавливает свою функцию, в результате чего восстанавливается фенотип. Такое восстановление (полное или частичное) фенотипа вопреки сохранению первоночального генетического повреждения (мутации) получило название супрессии, а такие обратные мутации назвали супрессор-ными (внегенными). Как правило, супрессии происходят в результате мутаций генов, кодирующих синтез тРНК и рибосом.
Большинство генов довольно устойчива по отношению к мутациям, однако известны гены, которые мутируют очень часто. Как правило, мутантный фенотип организмов является одинаковым, независимо от того, наследуется ли генная мутация от отца или матери. Тем не менее небольшое число генов у человека и мышей экспрессируется дифференциально, причем в зависимости от источника наследования мутантного гена (от отца или матери). Во всех случаях один из родительских аллелей полностью инактиви-руется, тогда как другой аллель является активным. Напоминая инактивацию Х-хромосом, это явление получило название генетического импринтинга. Следовательно, генетический импринтинг представляет собой явление неэквивалентности генных аллелей, активность которых зависит от родительского происхождения. Например, патологический синдром Прадела-Вилли у человека обязан мутации, которая встречается только в отцовском хромосомном гомологе, а синдром Ангельмана у человека возникает в результате мутации в материнском хромосомном гомологе.
Хромосомные мутации связаны с изменениями числа и структуры хромосом. Изменения в числе хромосом определяются добавлением или уменьшением всего набора хромосом, ведущим к полиплоидии или гаплоидии (соответственно), часто называемые геномными мутациями, а также добавлением или удалением одной или больше хромосом из набора, что ведет к гетероплоидии или анеуплоидии (моносомии, трисомии и др. полисемии), тогда как изменения в структуре хромосом определяются перестройками (аберрациями) их структуры.
Полиплоидия — это хромосомная мутация в виде увеличения числа полных гаплоидых наборов хромосом. Известны триплоидия (Зп), тетраплоидия (4п), пентаплоидия (5п) и т. д. Наиболее часто полиплоидия встречается у растений, поскольку для них характерны гермафродитизм и апомиксис. Почти третья часть всех видов диких цветковых растений представлена полиплоидами. Типичными полиплоидами являются виды пшеницы, у которых соматические числа 2n = 14,28 и 42, при основном гаметном числе n = 7, картофель, табак, белый клевер, люцерна и другие растения. Родственные виды, наборы хромосом которых представляют ряд возрастающего увеличения (кратного) основного числа хромосом, составляют полиплоидные ряды.
Полиплоидные растения с нечетными наборами хромосом характеризуются пониженной плодовитостью и снижением уровня количественных признаков. Напротив, для растений-полиплоидов с четными наборами хромосом характерен повышенный уровень количественных признаков.
Полиплоиды, происходящие от диплоидных организмов аналогичного вида, называют автополиплоидами. Примером автопо-липлоидов служат растения мягкой пшеницы, у которых 2п == 42 и которые являются гексаплоидами. Искусственные полиплоиды, полученные из гибридов диплоидных растений, относящихся к далеко отстоящим друг от друга видам, называют аллополипло-идами.
В природе встречаются как автополиплоиды, так и аллополип-лоиды, однако географическое распределение полиплоидов обычно отличается от распределения их диплоидиых «родственников». Например, флора о. Шпицберген содержит очень высокий процент полиплоидных рядов, тогда как в других местах количество их меньше по сравнению с диплоидами.
Полиплоидия растений имеет хозяйственную ценность (повышенные размеры плодов, большая сахаристость, лучшая сохранность и др.). По этой причине полиплоиды используют в селекционной работе для получения новых сортов сельскохозяйственных растений.
У животных полиплоидия очень редка. Она встречается у земляных червей, размножающихся партеногенетически (обнаружены полиплоидные ряды с основными числами 11, 16, 17, 18 и 19 хромосом), у некоторых членистоногих, рыб и земноводных. В частности, она описана у морских креветок Artemia salivana. Женские особи саламандр отдельных видов, которые имеют крупные эритроциты с ядрами, продуцируют триплоидные личинки с 42 хромосомами, тогда как саламандры с малыми ядерными эритроцитами продуцируют диплоидные личинки с 28 хромосомами. Все тихоокеанские лососевые рыбы являются полиплоидами.
Сирийский хомячок (Mesocricetius aurantus), у которого 2 n = 44, является аллополиплоидом, возникшим в результате естественной гибридизации между европейским хомяком (Cricetus cricetus, 2n == 22) и хомяком, принадлежащим к одному из азиатских видов (Cricetus griseus, 2n = 24).
Полиплоидия встречается также у человека в пренатальном периоде развития. В частности известно много сообщений об обнаружении триплоидии и тетраплоидии в клетках абортусов. Имеющиеся данные позволяют считать, что триплоидия встречается у 20% абортусов, а тетраплоидия — у 6% абортусов.
Редкость полиплоидии у раздельнополых животных, по-видимому, определяется тем, что она нарушает нормальные соотношения аутосом и половых хромосом в зиготах.
Кроме авто- и аллополиплоидии, в соматических клетках ряда многоклеточных организмов установлена эндополиплоидия, характеризующаяся увеличением числа хромосом в покоящемся ядре (при отсутствии митоза). От полиплоидии следует отличать псевдополиплоидию отдельных растений и насекомых, возникающую в результате однократного или многократного деления компонентов генома, когда центромеры имеют диффузный характер.
Гаплоидия — это мутация в виде уменьшения всего набора хромосом. Она найдена в основном также у растений. Известны гапло-иды свыше 800 видов растений, включая такие культуры, как пшеница и кукуруза. У животных она очень редка, а у человека совсем неизвестна.
Мутации в виде нарушений нормального количества хромосом из-за добавления или удаления одной или более хромосом в какой-то хромосомной паре называют гетероплоидией или анеуп-лоидией. Среди гетероплоидов известны полисемии, когда какая-то пара хромосом становится большей на один (трипликатом) и более гомологов, моносомии, когда какая-либо хромосомная пара теряет один гомолог, и нулесомии, когда теряется вся хромосомная пара. Эти мутации широко распространены как у животных, так и у растений. В частности трисомии и моносомии обнаружены у человека, собак и других животных, а также у многих плодовых, зерновых и овощных растений. Часто трисомии, как и моносомии, обнаруживают в клетках абортированных эмбрионов или плодов человека. Гетероплоидия сопровождается многими фенотипическими эффектами, которые, например, у человека неблагоприятны для его здоровья.
Мутации, поражающие структуру хромосом, называют хромосомными перестройками или чаще аберрациями. Эти мутации классифицируют на внутрихромосомные и межхромосомные перестройки. Они возникают как у животных, включая человека, так и у растений.
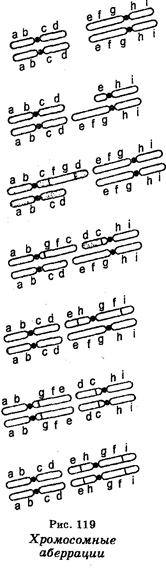 Внутрихромосомными перестройками являются делении (нехватки, дефишенсы), дубликации, инверсии, тогда как межхромосомные перестройки представлены транслокациями (рис. 119). Внутрихромосомные перестройки в зависимости от «разлома» плеч хромосом бывают парацентрическими («разлом» затрагивает одно хромосомное плечо) и перицентрическими («разлом» проходит по обе стороны центромеры, т. е. захватывает оба плеча). Межхромосомные перестройки обычно захватывают две или более негомологичные хромосомы. Степень изменений бывает различной и часто доходит до изменения группы сцепления.
Внутрихромосомными перестройками являются делении (нехватки, дефишенсы), дубликации, инверсии, тогда как межхромосомные перестройки представлены транслокациями (рис. 119). Внутрихромосомные перестройки в зависимости от «разлома» плеч хромосом бывают парацентрическими («разлом» затрагивает одно хромосомное плечо) и перицентрическими («разлом» проходит по обе стороны центромеры, т. е. захватывает оба плеча). Межхромосомные перестройки обычно захватывают две или более негомологичные хромосомы. Степень изменений бывает различной и часто доходит до изменения группы сцепления.
Делеции представляют собой потери сегмента хромосомы, несущего один или несколько генов. Они являются наиболее частой и опасной для человека формой генетических макроповреждений. Большие делеции заключаются в потере одного или нескольких генов или даже блоков генов. У гаплоидных организмов крупные делеции детальны. Эффект делеции у диплоидных организмов зависит от количества делегированных генов, количественных потребностей в продуктах пораженных генов, позиции генов среди функционально координированных групп и других факторов. Гомозиготные делеции для диплоидных клеток или организмов детальны.
Дупликации (добавления) представляют собой добавления (удлинения) какого-либо сегмента хромосомы, несущего один или несколько генов, в результате того, что один и тот же сегмент хромосомы может быть повторен в процессе биосинтеза несколько раз. Этот повтор может быть малым, касаясь одиночного гена, или большим, затрагивая большое количество генов. Дупликации часто безвредны для их носителей. Предполагают, что они способствуют формированию полигенов или являются способом введения новых генов в геномы. Некоторые дуп-ликации, однако, вредны и даже детальны (см. гл. IX).
Инверсии заключаются в поворотах на 180° сегментов, освобождающихся в результате парных разрывов в хромосомах. Если инвертированный сегмент не содержит центромеру, эту мутацию называют парацентрической инверсией, если же такой сегмент содержит центромеру, такую мутацию называют перицентрической. Инверсии оказывают влияние на мейоз, что приводит к пониженной фертильности гибридов. Описаны отдельные наследственные аномалии, вызываемые этой мутацией (см. гл. XIII).
Транслокация — это обмен частями (сегментами) гомологич-ной и негомологичной хромосом, образованными разрывами по длине последних. Транслоцируемые сегменты могут иметь разные размеры — от небольших до значительных.
Хромосомные перестройки, как и гетероплоидия, также сопровождаются различными фенотипическими эффектами. У человека они ведут к нарушению здоровья. У растений они ведут к изменению их продуктивности.
Однако, обсуждая значение хромосомных перестроек, нельзя не заметить, что для отдельных организмов они безразличны. Например, у пионов, дурмана и других растений присутствие в гено-ме транслокаций является нормальным явлением.
В зависимости от происхождения различают спонтанные и индуцированные генные и хромосомные мутации, которые возникают у организмов независимо от уровня их организации.
Спонтанными называют те мутации, которые возникают у организмов в нормальных (природных) условиях на первый взгляд без видимых причин, тогда как индуцированными называют те мутации, которые возникают в результате обработки клеток (организмов) мутагенными факторами. Главное отличие спонтанных мутаций от индуцированных заключается в том, что мутация может возникнуть в любой период индивидуального развития. Что касается случайного характера мутаций в пространстве, то это означает, что спонтанная мутация произвольно может поразить любую хромосому или ген.
Длительное время считали, что спонтанные мутации являются беспричинными, однако теперь по этому вопросу существуют другие представления, сводящиеся к тому, что спонтанные мутации не являются беспричинными, что они являются результатом естественных процессов, протекающих в клетках. Они возникают в условиях природного радиоактивного фона Земли в виде космического излучения, радиоактивных элементов на поверхности Земли, радионуклидов, инкорпорированных в клетки организмов, которые вызывают эти мутации или в результате ошибок репликации ДНК. Факторы естественного радиоактивного фона Земли вызывают изменения в последовательности оснований или повреждения оснований подобно тому, как это имеет место в случае индуцированных мутаций (см. ниже).
Ошибки репликации ДНК заключаются в том, что какой-либо нуклеотид ошибочно включается в ДНК при ее репликации и при наличии недостатка в «редактирующем» механизме, что исключает коррекцию ошибок. Ошибки в репликации ДНК, т. е. «неаккуратность» ДНК-полимеразы оказывают влияние на частоту спонтанных мутаций. Ошибки репликации ДНК могут быть также связаны с химической нестабильностью нуклеотидов, сопровождающейся изменением спаривающей способности азотистых оснований. Например, цитозин может быть дезаминирован в урацил, который затем распознается ДНК-полимеразой как тимин в течение репликации ДНК.
Определение частот спонтанных мутаций организмов разных видов проводят с помощью разных способов, один из которых связан с определением частоты таких мутаций на репликацию пары азотистых оснований в молекулах ДНК. Этот способ оказался предпочтительным в случае бактерий и других низших организмов.
Определенные к настоящему времени частоты мутаций на репликацию пары оснований и общие частоты генных мутаций у разных организмов показаны в табл. 13.
В случае человека частоты спонтанных мутаций определяют измерением прямых мутаций в пределах разных генов, которые очень чувствительны к мутациям независимо от того, являются ли условия для организмов ограничивающими или селективными.
Рассмотрим конкретный пример определения частоты спонтанных мутаций, например, среди родившихся за один год 242 257 детей 7 оказались больными ахондоплазией. Следовательно, 7: 242 257´1: 2 (два аллеля на зиготу) = 1,4´10-6. Таким образом частота ахондоплазии составляет 1,4 х 10-5. Одни гены вообще устойчивы к спонтанным мутациям, другие спонтанно мутируют чаще, третьи так часто, что их носители являются мозаиками мутантных (мутировавших) и немутантных (немутировавших) генов.
Средние частоты мутаций по многим генам у человека и домашних животных составляют примерно 1´10-9, что значительно выше частоты мутаций микроорганизмов. Больше того, между частотами спонтанных мутаций по разным генам человека или домашних млекопитающих существуют значительные различия, достигающие 100 раз, а то и более. Подлинные причины этих различий неизвестны, хотя для их объяснения и предложено несколько гипотез. Одна из них заключается в том, что наиболее чувствительны к мутациям гены больших размеров, поскольку в них содержится много азотистых оснований и существует большая вероятность мутации отдельных из них. По другой гипотезе наиболее чувствительными к мутациям являются гены, располагающиеся в районах хромосом, являющихся «горячими» точками.
Таблица 13
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 552; Нарушение авторских прав?; Мы поможем в написании вашей работы!