КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
ЛЕКЦИЯ №9
|
|
|
|
Произвольные движения человека – это сознательно регулируемые движения, которые осуществляются в отличие от непроизвольных с определенной целью.
Механизмы инициации движения
Центральные аппараты и механизмы управления движениями
Формирование двигательных программ и механизм управления движением
РЕГУЛЯЦИЯ ПРОИЗВОЛЬНЫХ ДВИЖЕНИЙ
ЛЕКЦИЯ №8
Взаимодействие человека и животных с окружающей средой может осуществляться только через движение. Существует два вида двигательных функций: поддержание положения тела (позы) и собственно движения. Среди собственно движений следует различать элементарные движения и действия. Последние – действия, представляют собой совокупность элементарных движений, объединенных единой целью в некоторую функциональную систему. Каждое конкретное действие включено в состав поведенческого акта. Оно является шагом к удовлетворению той потребности, которая вызвала данное поведение, и решает свою промежуточную задачу.
Произвольные двигательные акты связаны с функцией пирамидального и экстрапирамидального трактов, связывающих кору головного мозга и подкорковые структуры со спинальным моторным механизмом (мотонейронами спинного мозга).
Впервые наличие прямых эфферентных и обратных афферентных связей между нервными структурами спинного мозга и мышечным аппаратом была показана в работе Чарльза Белла, который описал нервный круг: один нерв несет импульсы мозга к мышце, другой приносит ощущения состояния мышцы к мозгу.
Позже И.М. Сеченов выделил понятие «темное мышечное чувство», которое по его убеждению порождает представление о скорости движения и отношениях предметов в пространстве. Рецепторный аппарат мышц, сухожилий и суставов, образующих «мышечное чувство» Ч.С. Шеррингтон назвал проприорецепторами. Шеррингтон считал, что главная функция проприорецепторов – давать информацию о собственных движениях организма. В настоящее время выделено два типа проприорецепторов: рецепторы растяжения и рецепторов натяжения мышечных волокон. В эксперименте в 1909 году И.П. Павлов установил, что моторная кора получает сенсорные сигналы от рецепторов мышц и суставов и ввел понятие двигательного анализатора. В результате чего моторная кора стала рассматриваться как центральный аппарат построения движений.
Обобщив перечисленные факты, П.К. Анохин применил понятие обратная связь или обратная афферентация. Таким образом, основной принцип в механизме координации двигательных реакций является циклическая нервная связь и кольцевая структура управления или рефлекторное кольцо по Н.А. Бернштейну.
Однако не все двигательные акты осуществляются по кольцевому механизму – движения, базирующиеся на врожденных координациях, в меньшей степени требуют обратной связи, в то время как приобретаемые движения всецело зависят от соматической афферентации с двигательного аппарата. Движения, осуществляющиеся без помощи проприоцептивной обратной связи, выполняются по жесткой программе. Но даже эти движения требуют сличения с определенным сенсорным образом. П.К. Анохин для обозначения механизма сличения выделил в функциональной системе двигательного акта специальный блок сличения, который назвал акцептор результата действия.
В управлении движением различают стратегию и тактику. Основу стратегии движения определяет конкретная мотивация (биологическая, социальная и др.). Именно на ее основе определяется цель поведения, т. е. то, что должно быть достигнуто. В структуре поведенческого акта цель закодирована в акцепторе результатов действия. В отношении движения это выглядит как формирование двигательной задачи, т. е. того, что следует делать. Под тактикой понимают конкретный план движений, т. е. то, как будет достигнута цель поведения, с помощью каких двигательных ресурсов, способов действия.
Таким образом, можно выделить два механизма управления движением:
- посредством центральных моторных программ;
- с помощью обратной афферентации.
Впервые представление о центральных моторных программах было дано в работе Ч. Шеррингтона, который занимался изучением функций мотонейронов спинного мозга собак при осуществлении двигательных актов ходьбы и чесания. Производя перерезки участков спинного мозга, он показал, что механизмы, ответственные за некоторые двигательные акты локализованы в спинном мозге. Через несколько месяцев после перерезки у собак удавалось вызвать чесательный рефлекс на механическое раздражение кожи. На основании чего была заложена концепция о «запускаемом движении» с помощью центральных программ. Позже было показано, что ходьба животного – перемещение в окружающей среде при помощи координированных движений конечностей – это также запрограммирована на уровне спинного мозга.
Программы центральных моторных программ широко представлены в различных структурах мозга и ЦНС хранит как врожденные, так и приобретенные программы выработанных действий. Например, дыхательные, глотательные движения управляются врожденными программами, а профессиональные, спортивные и иные навыки осуществляются по программам, формирующимся в течение жизни индивида. При определенном опыте человека эти программы выполняются автоматически, а обратная афферентация перестает играть существенную роль. Обратная афферентация становится нужна при переделке – корригировании (корректировке) навыка.
У позвоночных центральные программы запускаются управляющими центрами, а у беспозвоночных, как правило, командными нейронами, впервые выделенными в 1968 г. К. Вирсма. Однако сигнал от командного нейрона или управляющего центра играет лишь пусковую роль. Он не несет информации о том, как построить соответствующее движение. Структура движения заложена во взаимосвязях мотонейронов, с которыми он связан.
Рассмотрим несколько примеров поведения у низших животных:
- рефлекс бегства у рака, легко вызываемый тактильным, вибрационным или звуковым раздражением, реализуется через возбуждение одного гигантского командного нейрона, связанного с тремя двигательными мотонейронами;
- в ответ на стимуляцию рыба уплывает, совершая быстрые и повторяющиеся движения хвостом и головой; этот двигательный акт запускается парой командных нейронов – маутнеровских клеток, расположенных в продолговатом мозгу;
- у таракана есть командный нейрон, запускающий бег насекомого, если его лапы касаются опоры, если же лапы находятся в воздухе запускается команда полета.
Существует два типа командных нейронов: одни лишь запускают двигательную программу, не участвуя в ее дальнейшем осуществлении, другие поддерживают или видоизменяют двигательную программу. Первый тип нейронов получил название триггеры. Второй тип нейронов получил название воротных нейронов.
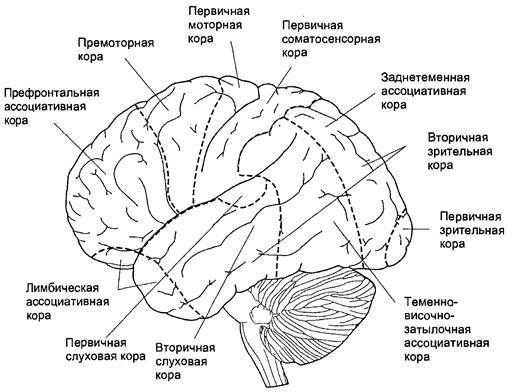
Рисунок 3 – Схема двигательной коры большого мозга
Ведущая роль в построении новых моторных программ у высших животных и человека принадлежит передним отделам коры больших полушарий – премоторной и префронтальной коре. Роль передних отделов коры больших полушарий показана в работах А.С. Батуева, проводившего эксперименты с использованием микроэлектородов, регистрирующих активность отдельных нейронов у обезьян.
В экспериментах были обнаружены три группы нейронов, последовательно вовлекаемых в процесс осуществления условнорефлекторного двигательного навыка. Все три популяции клеток были обнаружены в теменной и особенно в лобной коре.
Первая группа клеток реагирует на условные сигналы, поэтому их отнесли к сенсорным нейронам. Вторая группа клеток реагирует на время действия сигнала, а третья группа в пусковой период. Считают, что сенсорные нейроны первой группы передают информацию второй группе, которые принадлежат кратковременной памяти. Третья же группа – это нейроны моторных программ, они получают информацию от нейронов памяти и запускают хорошо отработанную двигательную реакцию.
Многие исследователи считают, что хранение части подобных моторных программ осуществляется структурами мозжечка. Кроме того, в процессе индивидуальной жизни мозжечок обучается различным программам движения и сохраняет их. В этом процессе важную роль играют клетки Пуркинье. Мозжечок получает информацию о планируемом корой большого мозга движении, вносит коррекцию в план будущего движения и посылает ее в кору большого мозга. Быстрые мелкие точные движения осуществляются без сличения корой с импульсами от проприорецепторов. Мозжечок обучается также корригировать (изменять) выполняемое движение, играет важную роль в регуляции тонуса мышц, поддержании позы, координации движения.
В качестве другой центральной структуры, связанной с хранением центральных моторных программ рассматриваются базальные ганглии. Базальные ганглии имеют отношение к хранению программ врожденных двигательных актов и быстрых стереотипных движений. Врожденные программы имеются также в спинном мозге (шагательный рефлекс) и в стволе мозга (дыхания, глотания, выпрямительный рефлекс – восстановление нарушенной позы, ориентировочный рефлекс).
Инициация движения происходит следующим образом. В моторной коре строится конечный и конкретный вариант моторного управления движением. Моторная кора использует оба принципа управления: контроль через систему обратной афферетации и через механизм центрального программирования. Это достигается тем, что к ней сходятся сигналы от мышечной активности, от сенсомоторной, зрительной и других отделов коры, которые и используются для моторного контроля коррекции движения. Кроме того, к моторной коре приходят сигналы, связанные с программированием движения из передних отделов коры и подкорки.
Моторная кора получает также информацию из структур, имеющих отношение к хранению двигательных программ. Так, в опытах с регистрацией нейронной активности у обезьян было показано, что при выполнении ими заученного движения активность нейронов зубчатого ядра мозжечка на 10 мс опережает изменение активности нейрона в моторной коре, которое предшествует появлению мышечного движения.
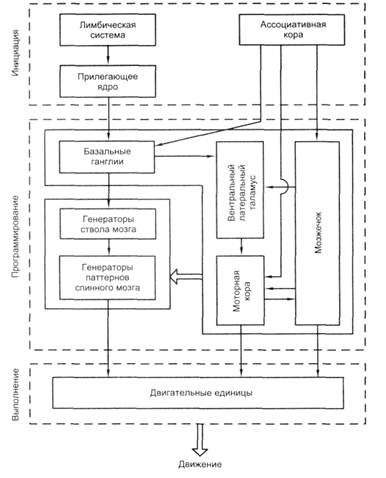
Рисунок 2 - Схема инициации двигательного акта (по Г. Могенсону, 1977, с изменениями)
В моторную кору поступают также сигналы от базальных ганглиев – структура, которая ответственна за хранение главным образом двигательных программ врожденного поведения (пищевого, питьевого и др.), стереотипных движений. Клетки базальных ганглиев, так же как и мозжечка, разряжаются до начала движений, совершаемых животными в ответ на сигнал за 50 – 150 мс. В хвостатом ядре были найдены нейроны, у которых возбуждение опережает условную двигательную пищевую и оборонительную реакции. Эти реакции у нейронов появляются с выработкой условного рефлекса и исчезают с его утешением. Сигналы от базальных ганглиев и мозжечка распространяются не только вверх к коре, но и вниз, по нисходящему пути к спинному мозгу (так называемый экстрапирамидный путь).
Согласно работам шведского нейрофизиолога Г. Могенсона, все процессы управления движением включают три блока (рассмотрим рисунок 2) и соответствующие им три фазы:
1) блок инициации движения, включающий лимбическую систему с прилегающим ядром (n. accumbens), и ассоциативную кору;
2) блок программирования движения, включающий мозжечок, базальные ганглии, моторную кору, таламус (как посредник между ними), а также спинальные и стволовые генераторы;
3) исполнительный блок, охватывающий мотонейроны и двигательные единицы.
Таким образом, произвольные движения, так же как и непроизвольные, являются результатом сопряженной работы лимбической системы и ассоциативной коры. Двигательная система работает как многоуровневая. На каждом уровне имеется своя «ведущая афферентация» и собственный тип регулируемых движений. Однако произвольные движения без пирамидной системы осуществляться не могут, а непроизвольные – могут. Непроизвольные движения запускаются посредством возбуждения экстеро- и проприорецепторов, могут осуществляться с помощью любого уровня ЦНС, в том числе и на уровне спинного мозга. Произвольные движения инициируются ассоциативной корой и лимбической системой с помощью механизмов памяти.
|
|
|
|
|
Дата добавления: 2013-12-12; Просмотров: 956; Нарушение авторских прав?; Мы поможем в написании вашей работы!