КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Память – это способность организма приобретать, сохранять и воспроизводить в сознании информацию и навыки
|
|
|
|
ПСИХОФИЗИОЛОГИЯ ПАМЯТИ
Лекция №12
1 Физиологические теории памяти
2 Классификация видов памяти. Кратковременная и промежуточная память
3 Долговременная память и молекулярные механизмы ее консолидации
Основу адаптивного (индивидуального) поведения составляют два процесса - обучение и память. В нейрологической памяти выделяют генотипическую (врожденную) память, которая обусловливает становление безусловных рефлексов, инстинктов, импринтинга, и фенотипическую память, мозговые механизмы которой обеспечивают обработку и хранение информации, приобретаемой живым существом в процессе индивидуального развития. Современная наука дает следующее определение памяти:
Обучение и память считают неотделимыми процессами. Обучение обеспечивает постоянное пополнение и изменение знаний, приобретение новых навыков. С физиологической точки зрения научение - это результат совпадения двух сознательных или бессознательных процессов в головном мозге. В отличие от обучения процессы памяти ответственны не только за усвоение (фиксацию) информации, ее сохранение, но и включают механизм воспроизведения (извлечения) информации. Благодаря механизму воспроизведения обеспечивается доступ и использование хранящейся информации.
Исследование механизма индивидуальной памяти, по существу, представляет собой область изучения механизмов различных видов обучения. Память и обучение имеют общую особенность — необходимость повторения. Как научение основано на многократном сочетании одних и тех же стимулов, так и для образования следа памяти требуется повторение одной и той же информации.
В современной нейробиологии и психофизиологии существует целый ряд теорий и моделей, объясняющих разные стороны функционирования памяти.
Теория Дональда Хебба. Первые исследования физиологических основ памяти связаны с именем Дональда Хебба. В 40-е гг. он ввел понятия кратковременной и долговременной памяти и предложил теорию, объясняющую их нейрофизиологическую природу. По Хеббу, кратковременная память — это процесс, обусловленный повторным возбуждением импульсной активности в замкнутых цепях нейронов, не сопровождающийся морфологическими изменениями. Долговременная память, напротив, базируется на структурных изменениях, возникающих в результате модификации межклеточных контактов — синапсов. Хебб полагал, что эти структурные изменения связаны с повторной активацией (по его определению — "повторяющейся реверберацией возбуждения") замкнутых нейронных цепей.
Повторное возбуждение нейронов, образующих такую цепь, приводит к тому, что в них возникают долговременные изменения, связанные с ростом синаптических соединений и увеличением площади их контакта между пресинаптическим аксоном и постсинаптической клеточной мембраной. После установления таких связей эти нейроны образуют клеточный ансамбль, и любое возбуждение хотя бы одного относящегося к нему нейрона, приводит в возбуждение весь ансамбль. Это и есть нейрональный механизм хранения и извлечения информации из памяти.
Основные структурные изменения, согласно Хеббу, происходят в синапсах в результате процессов их роста или метаболических изменений, усиливающих воздействие каждого нейрона на следующий нейрон. Достоинство этой теории в том, что она толкует память не как статическую запись или продукт изменений в одной или нескольких нервных клетках, а как процесс взаимодействия многих нейронов на основе соответствующих структурных изменений.
Синаптическая теория утверждает, что при прохождении импульса через определенную группу нейронов возникают стойкие изменения синаптической проводимости в пределах определенного нейронного ансамбля.
Один из наиболее авторитетных исследователей нейробиологических основ памяти, Стивен Роуз [5] подчеркивает: при усвоении нового опыта, необходимого для достижения каких-либо целей, происходят изменения в определенных клетках нервной системы. Эти изменения, выявляемые морфологическими методами с помощью световой или электронной микроскопии, представляют собой стойкие модификации структуры нейронов и их синаптических связей.
Г. Линч и М. Бодри (1984) предложили следующую гипотезу. Повторная импульсация в нейроне, связанная с процессом запоминания, предположительно, сопровождается увеличением концентрации кальция в постсинаптической мембране, что приводит к расщеплению одного из ее белков. В результате этого освобождаются замаскированные и ранее неактивные белковые рецепторы (глутаматрецепторы). За счет увеличения числа этих рецепторов возникает состояние повышенной проводимости синапса, которое может сохраняться до 5-6 суток.
Эти процессы тесно связаны с увеличением диаметра и усилением активности так называемого аксошипикового синапса — наиболее пластичного контакта между нейронами. Одновременно с этим образуются новые шипики на дендритах, а также увеличиваются число и величина синапсов. Таким образом, экспериментально показаны морфологические изменения, сопровождающие формирование следа памяти.
Реверберационная теория. Основания теории были выдвинуты известным нейрофизиологом Лоренто де Но. Теория базировалась на существовании в структурах мозга замкнутых нейронных цепей. Известно, что аксоны нервных клеток соприкасаются не только с дендритами других клеток, но могут и возвращаться обратно к телу своей же клетки. Благодаря такой структуре нервных контактов, появляется возможность циркуляции нервного импульса по реверберирующим (постепенно затухающим) кругам возбуждения разной сложности. В результате возникающий в клетке разряд возвращается к ней либо сразу, либо через промежуточную цепь нейронов и поддерживает в ней возбуждение. Эти стойкие круги реверберирующего возбуждения не выходят за пределы определенной совокупности нервных клеток и рассматриваются как физиологический субстрат сохранения энграмм памяти. Именно в реверберационном круге возбуждения происходит переход из кратковременной в долговременную память.
С работой Лоренто де Но связана гипотеза А.С. Батуева о двух нейронных системах, обеспечивающих оперативную память. Одна система, включающая "нейроны памяти", работает на эстафетно-реверберационном принципе передачи информации, когда отдельные группы нейронов памяти вовлекаются друг за другом, представляя собой своеобразные "нейронные ловушки", поскольку возбуждение в них циркулирует в течение 1,5-2 с. Другая система обеспечивает надежность переходных процессов: переключение информации с "сенсорных" нейронов на "нейроны памяти" и далее на нейроны "моторных программ" и т.д. Их взаимодействие позволяет эффективно запоминать текущую информацию.
Однако реверберационная теория не дает ответа на ряд вопросов. В частности, она не объясняет причину возврата памяти после электрошоковых воздействий, когда, согласно этой теории, в подобных случаях возврата памяти не должно быть.
Как было сказано выше в нейрологической памяти выделяют генотипическую (врожденную) память, которая обусловливает сохранение инстинктов, импринтинга, и фенотипическую память, мозговые механизмы которой обеспечивают обработку и хранение информации, приобретаемой животным организмом в процессе индивидуального развития.
Кроме того память различают по формам восприятия информации:
· логически-смысловая
· чувственно-образная
Последняя подразделяется на:
- модально-специфические виды – зрительную, слуховую, моторную
- по уровням усвоения (воспроизводящая и облегчающая),
- по длительности хранения информации.
Онтогенетическую память составляют условные рефлексы и другие формы приобретенной информации.
По длительности хранения информации различные авторы выделяют несколько вариантов памяти, причем везде первым этапом отмечается сенсорная (по мнению некоторых авторов – иконическая) память – доли секунды.
а) первичная память – несколько секунд, вторичная – от нескольких минут до нескольких лет, третичная – на протяжении всей жизни;
б) кратковременная память – десятки секунд, кратковременная (первичная) – десятки секунд, кратковременная (вторичная) – от нескольких минут до нескольких лет,
в) долговременная – долговременная – более минуты и на протяжении всей жизни.
Кратковременная память. Этот вид памяти формируется на основе непосредственного сенсорного отпечатка внешнего мира. Наличие кратковременной памяти было доказано в опытах немецкого психолога Германа Эббингауса (1885), проводившего оценку эффективности воспроизведения человеком рядов случайных цифр, букв, символов. Г. Эббингаус впервые установил, что случайные последовательности цифр, букв, символов после однократного чтения, прослушивания можно воспроизвести без ошибок в строго ограниченном количестве единиц. Память, обеспечивающая удержание и воспроизведение оперативной информации, получила название кратковременной. Объем кратковременной памяти составляет примерно 7 ± 2 единицы. Основным свойством данной разновидности памяти является ее непродолжительность. Длительность хранения информации в кратковременной памяти – секунды, минуты.
Единой теории механизма кратковременной памяти до сих пор не существует. Однако ясно, что запоминание любой информации начинается с электрофизиологических процессов в нейронных сетях головного мозга (возникновение ВПСП, ПД, выделение различных медиаторов в синаптическом аппарате мозга). Поэтому можно утверждать, что в основе механизма кратковременной памяти лежит импульсная активность нейронов и, в частности, циркуляция возбуждения по замкнутым нейронным цепям.
Большую роль для выяснения механизмов памяти сыграло применение в лечебных целях электрошока – сильного электрического воздействия на головной мозг. Выяснилось, что после электрошока, так же как и после механической травмы мозга в результате сотрясения, наркоза, ушиба, наблюдается ретроградная амнезия: человек полностью забывает о событиях, предшествовавших электрическому или механическому воздействию на мозг. По длительности ретроградной амнезии можно получить представление о времени консолидации памяти (от лат. consolidate -упрочение, укрепление, объединение), т.е. времени перехода кратковременной памяти в долговременную. Консолидация памяти начинается обычно через несколько минут, иногда через несколько десятков минут (в опытах на крысах, у которых после выработки условного рефлекса вызывали электрошок, приобретенный навык сохранялся, если электрошок проводился через 10 мин и более после начала обучения, при более раннем воздействии на мозг – навык исчезал).
Наблюдения за развитием состояния ретроградной амнезии у человека привели к формулировке одной из первых гипотез о механизме кратковременной памяти – гипотезе реверберации (циркуляции) возбуждения по замкнутым цепочкам нейронов. Считают, что в замкнутых нейрональных цепочках циркуляция длится минутами, сохраняя информацию в виде последовательности импульсов, передающихся от нейрона к нейрону (рис.). Известно, что пока циркуляция продолжается, сохраняется нейрональный след о воздействии того или иного раздражителя на организм в прошлом. С позиций гипотезы реверберации легко объясняется феномен ретроградной амнезии. Таким образом, развитие амнезии при травмах головного мозга, при электрошоке возникает вследствие грубого нарушения ритмической активности в цепочках взаимосвязанных нейронов.
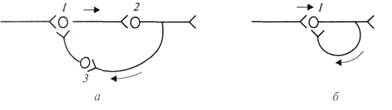
Рис. Циркуляция возбуждения в замкнутых нейронных цепях (по Лоренто де-Но – а и по И. С. Беритову – б):1, 2, 3 – возбуждающие нейроны
Запоминание начинается с электрофизиологических процессов (изменение ВПСП, ПД), поэтому кратковременную память по механизму ее происхождения можно назвать электрофизиологической памятью.
Промежуточная память – это процесс перевода кратковременной памяти в долговременную память (консолидация памяти), который продолжается несколько часов.
Следы кратковременной памяти становятся устойчивыми через 4 ч. Таким образом, длительность промежуточной памяти от нескольких минут до 4 ч.
Формирование промежуточной памяти связано не только с электрофизиологическими процессами, но и цепью биохимических реакции, активирующих синтез медиаторов, рецепторов ионных каналов. Поэтому данный период переработки поступившей информации и ее запоминание следует назвать нейрохимической памятью. В этот период экстремальные воздействия (наркоз, сотрясение мозга и др.) еще способны стереть память.
Основой долговременной памяти являются структурные изменения в нейронах, длящиеся или сохраняющиеся на протяжении часы, дни и всей жизни при повторении информации. Ее объем практически безграничен. Долговременная память по своему механизму качественно отличается от кратковременной и промежуточной памяти, так как не нарушается при таких экстремальных воздействиях на мозг, как механическая травма, электрошок, наркоз и т.д. Долговременная память формируется с помощью кратковременной и промежуточной памяти, при этом важную роль играют синаптические процессы и метаболические изменения в нейронных сетях.
Синаптические процессы. Под влиянием обучения в ЦНС образуются новые синапсы, увеличиваются их размеры, количество, количество медиаторов, разрастаются дендриты, увеличивается число шипиков на них, количество коллатералей аксонов – все эти явления в течение месяца исчезают. Различные медиаторы могут оказывать разные эффекты в процессах усвоения и хранения информации. Серотонин, в частности, ускоряет обучение и удлиняет сохранение навыков при положительном эмоциональном подкреплении, например пищевом, и блокирует у животных выполнение оборонительных навыков. Норадреналин ускоряет обучение в условиях применения отрицательного подкрепления (электрокожного).
Важную роль в консолидации долговременной памяти и взаимодействии нейронов играют нейропептиды.Он находятся в пресинаптических терминалях в качестве сопутствующего нейромедиатора.
Например, вместе с норадреналином часто выделяются нейропептид γ и опиоидные пептиды. Дофамин часто выделяется окончаниями аксонов вместе с энкефалином; ацетилхолин – с вазоактивным интестинальным пептитодом (ВИП), энкефалином; серотонин – с веществом Р, тиреолиберином, холецистокинином (И.П. Ашмарин).
Нейропептиды оказывают пре- и постсинаптическое модулирующее действие. Пептид-спутник может значительно повысить взаимодействие между клетками. Например, ВИП усиливает сродство к ацетилхолину более чем в 10000 раз. Кроме того, пептиды выделяются нервными клетками не только в синаптическую щель, но и во внеклеточное пространство. Эффективный путь диффузии пептидов может составить 1 мм и более. Этого достаточно, чтобы охватить своим влиянием микроансамбль нейронов, изменяя его суммарную возбудимость, синхронизируя реактивность его нейронов, обеспечивая тем самым их включение в адаптивный поведенческий акт.
Образующийся ансамбль/мозаика функциональных групп нейронов, подвергнутых нейрохимическому воздействию пептидов, может быть одним из механизмов функционального объединения нервных клеток в ходе обучения, в консолидации памяти.
Важное значение для объяснения механизма консолидации долговременной памяти имеют структурные изменения нейронов, связанные с синтезом белков, участвующих в переводе информации из кратковременной в долговременную форму ее хранения.
Так установлено, что геном нервных клеток имеет относительно высокий по сравнению с другими тканями уровень функциональной активности. Характерным для нервных клеток является также прогрессирующее увеличение в них (в течение индивидуальной жизни) числа открытых для синтеза уникальных кодов ДНК, чего не происходит в тканях других органов. В частности, у эмбриона человека в возрасте 22 недель число активных генов в нервной клетке составляет 8,2 %, у взрослого человека эта величина достигает 24,6 %, а в некоторых зонах мозга – 38%, тогда как в мышцах с возрастом она не меняется. Транскрибируемость ДНК и ее синтез в нейронах увеличиваются при обучении животных и содержании их в условиях информационно обогащенной среды.
Известна молекулярная гипотеза памяти П.К. Анохина (1968), согласно которой биохимические процессы, протекающие на уровне протоплазмы, формируют динамические изменения генома нейрона, вызывая перестройку кода РНК, следствием чего является синтез адекватных для данной ситуации новых молекул белка, являющихся хранителем полученной информации. Таким образом, РНК необходима для обучения вообще.
Участие нейроспецифических белков в формировании следов памяти доказывается усилением их новообразования в процессе обучения и регистрации расстройства памяти при блокаде их синтеза. При угнетении механизмов, регулирующих синтез нейроспецифических белков, выработанные условные рефлексы при простых формах обучения сохраняются лишь на протяжении нескольких минут, иногда нескольких десятков минут. Затем в динамике этих условных рефлексов отмечаются явные расстройства, указывающие на нарушение процессов сохранения приобретенного навыка. Если синтез белка, обычно начинающийся в мозгу животного во время сеанса выработки рефлекса и длящийся много часов, блокировать, долговременного научения не произойдет.
Несколько сотен работ, опубликованных на эту тему, дают достаточно ясную картину: синтез белка в мозгу можно временно подавить на 80 – 90% без каких-либо грубых психических нарушений, если не считать памяти. Наиболее выраженная амнезия наблюдается и в случае подавления синтеза белка незадолго до сеанса обучения (и во время него уже не происходит). При этом даже спустя несколько недель повторения опытов усвоение остается неполным. Само по себе выполнение задания, а также ранее заученный материал при угнетении синтеза белка не страдают. Из этого следует, что он необходим лишь на критической стадии консолидации памяти непосредственно во время обучения или сразу после него. Для долговременного хранения информации синтез белка уже не нужен.
Подавление синтеза белка не влияет (по крайней мере, в опытах на животных) на кратковременную память. Это еще один важный аргумент, подтверждающий, что механизмы кратковременной и долговременной памяти различны.
В результате активации генома и синтеза специфических белков, в первую очередь в мембранах нервных клеток, в процессе обучения формируется структурно-функциональное объединение нейронов различных структур мозга, представляющее собой энграмму памяти.
Энграмма – это ансамбль нейронов и клеток глии, объединенных синаптическими механизмами. Такое динамическое объединение строится за счет определенных белковых молекул (адгезинов или коннектинов), которые встраиваются в специальные области мембран нейронов. Именно эти идентичные по молекулярным свойствам белковые молекулы увеличивают чувствительность нейронов к приему той информации, которая первично вызывала экспрессию этих белков.
Структурные основы воспоминания. Канадский нейрохирург У. Пенфилд первый показал, что стимуляция медиальных отделов височных долей коры полушарий у пациентов во время нейрохирургических операций вызывала детальные воспоминания событий, имевших место в отдаленном прошлом. При этом новые события оценивались пациентами как уже ранее пережитые.
При электрическом раздражении височных долей возникали живые картины из прошлого в их адекватной временной последовательности – занятия в школе или просмотренный кинофильм. Эти картины были очень ярки – больные переживали все подробности ранее происходивших событий, детали этих событий и соответствующие эмоциональные ощущения. Отмеченные картины четко воспроизводились при повторных стимуляциях медиальных отделов височной доли.
Вместе с тем, несмотря на приведенные примеры, демонстрирующие роль височной коры в извлечении опыта из памяти, следует иметь в виду, что память обусловлена деятельностью большого количества объединенных в структурно-функциональные ансамбли нейронов мозга, расположенных как в коре, особенно в лимбических структурах мозга, так и в подкорковых образованиях. Стимуляция височной коры только активирует эту корково-подкорковую энграмму памяти.
Процессы сохранения памяти в значительной степени связаны с функциями гиппокампа, амигдалы и височной коры. В хранении автоматизированных двигательных навыков существенная роль принадлежит структурам мозжечка.
Забывание. Процесс забывания связано с деятельностью структур гиппокампа и височной доли больших полушарий. Больные, у которых повреждены гиппокамп и височная доля, забывают приобретенные навыки очень быстро. У обезьян с разрушенным таламусом не отмечалось такого быстрого забывания, как после удаления гиппокампа. Быстрое забывание связано, по-видимому, с нарушением процесса консолидации памяти, т.е. переводом ее из кратковременной в долговременную память.
|
|
|
|
|
Дата добавления: 2013-12-12; Просмотров: 1368; Нарушение авторских прав?; Мы поможем в написании вашей работы!