КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Мутации у бактерий; мутагенные факторы
|
|
|
|
Фенотипическая изменчивость проявляется у подавляющего большинства особей в популяции, в то время как при мутационной изменчивости изменение генотипа происходит только у единичных клеток.
Понятие об адаптации микроорганизмов
Кроме генотипической, существует модификационная изменчивость, которая рассматривается как ответ на изменение условий окружающей среды и наблюдается до тех пор, пока действует фактор, вызывающий эти изменения. Модификационная изменчивость (ее называют еще фенотипической изменчивостью) проявляется на уровне фенотипа и не затрагивает генотип.
Модификация есть результат пластичности клеточного метаболизма, приводящего к фенотипическому проявлению «молчащих» генов в конкретных условиях. Таким образом, модификационные изменения имеют место в рамках неизменного клеточного генотипа.
Существует несколько проявлений модификационных изменений. Наиболее известны адаптивные модификации, т. е. ненаследственные изменения, полезные для организма и содействующие его выживанию в изменившихся условиях.
Причины адаптивных модификаций кроются в механизмах регуляции действия генов. Их примером может служить адаптация клеток бактерий E. coli к лактозе как новому субстрату: в этих условиях начинают синтезироваться индуцибельные ферменты, т. е. происходит фенотипическое проявление генов, «молчащих» при отсутствии лактозы в среде.
У ряда бактерий обнаружена универсальная адаптивная реакция в ответ на различные стрессовые воздействия (высокие и низкие температуры, резкий сдвиг рН и др.). В данном случае адаптивная реакция проявляется в интенсивном синтезе небольшой группы сходных белков, которые получили название белки теплового шока, а явление – синдром теплового шока. При стрессовых воздействиях на бактериальную клетку в ней ингибируется синтез обычных белков, но индуцируется синтез небольшой группы белков, функции которых заключаются в противодействии стрессовому воздействию путем защиты важнейших клеточных структур, в первую очередь нуклеоида и мембран. Считается, что адаптивные модификации расширяют возможности организма к выживанию и размножению в более широком диапазоне условий внешней среды. Возникающие модификации могут быть относительно стабильными, они могут сохраняться на протяжении нескольких поколений или, наоборот, очень лабильными.
Тем не менее, не все модификации можно рассматривать как адаптивные. При интенсивном воздействии многих агентов наблюдаются ненаследуемые изменения, случайные по отношению к вызвавшему их воздействию. Причины появления таких фенотипически измененных клеток связаны с ошибками процесса трансляции, вызванными этими агентами.
Значение адаптивных модификаций:
- вносят определенный вклад в процесс эволюции;
- расширяют возможности организма к выживанию и размножению в более широком диапазоне условий внешней среды. Возникающие в этих условиях наследственные изменения подхватываются естественным отбором и таким путем происходит более активное освоение новых экологических ниш и достигается более эффективная приспособляемость к ним.
Мутации – изменения, которые возникают в генетическом аппарате бактерий и передаются по наследству. Они бывают спонтанные и индуцированные. Мутации, возникающие в популяции бактерий без целенаправленного экспериментального вмешательства, называют спонтанными. Как правило, спонтанные мутации можно объяснить случайными ошибками при репликации ДНК. Возникают такие мутации довольно редко. В среднем частота спонтанных мутаций составляет 10–4–10–10.
Индуцированные мутации возникают с помощью воздействия тех или иных факторов – мутагенных агентов, которые существенно повышают частоту возникновения мутаций. Мутагенами могут быть химические, физические и биологические агенты, действующие на молекулу ДНК бактерий. К ним, например, относятся: УФ-лучи, ионизирующее излучение (физические агенты); азотистая кислота, нитрозогуанидин, аналоги азотистых оснований, некоторые антибиотики, акридиновые красители, сернистый иприт (химические агенты); транспозоны, IS-элементы, бактериофаг Mu (биологические агенты). Мутагенные агенты характеризуются неспецифичностью действия, т. е. используя какой-то мутаген, нельзя надеяться на выделение клеток с определенным типом или характером мутаций. Мутагены способны только повышать частоту возникновения мутаций.
По фенотипическим последствиям мутации подразделяют на прямые и обратные (реверсии). Мутации, которые приводят к утрате или изменению какой-то функции клетки, относятся к классу прямых, так как они вызывают появление у клеток другого фенотипа, который отличает их от бактерий дикого типа. Например, бактерии E. coli, способные в норме сбраживать лактозу (Lac+- фенотип), могут утрачивать данный признак, и поэтому мутация Lac+ → Lac– будет считаться прямой.
В результате обратной мутации у мутантного организма восстанавливается исходный (дикий) фенотип:
Lac– → Lac+ – обратная мутация (реверсия)
Обратные мутации бывают истинными (истинные реверсии) и вторичными. Об истинных обратных мутациях говорят лишь в тех случаях, когда в результате второй мутации восстанавливается исходный генотип. Однако эффект первой мутации может быть компенсирован мутацией в другой части этого же или расположенного рядом гена. Такие мутации называют вторичными реверсиями.
По фенотипическим проявлениям (характер проявления измененного признака) мутации подразделяют на:
1) морфологические, в результате которых изменяется ряд морфологических признаков (наличие капсулы, утрата жгутиков, изменение характерных особенностей колоний и др.);
2) биохимические, среди которых отмечены следующие:
• определяют зависимость от дополнительных факторов роста, или ауксотрофные;
• обеспечивают устойчивость к ингибиторам, антибиотикам, бактериоцинам, ядам или бактериофагам;
• связаны с чувствительностью к повышенной температуре (условно-летальные);
• изменяют способность использовать определенный субстрат или сбраживать какой-либо углевод;
• нарушают регуляцию или синтез ферментов катаболизма либо анаболизма;
• изменяют вирулентность клеток, их антигенные свойства, т. е. определяют характер взаимоотношений с другими организмами.
В соответствии с характером изменений в первичной структуре ДНК различают точковые и хромосомные мутации.
Точковые мутации – генные мутации, приводящие к замене, вставке или выпадению одной пары нуклеотидов. Для точковых мутаций с заменой оснований характерна высокая частота реверсии. Мутации такого рода могут быть двух типов:
• транзиции, в результате которых происходит замена пурина на другой пурин или же пиримидина на другой пиримидин (простая замена). Например, пара Г–Ц может быть заменена на пару А–Т, или наоборот. Это класс мутаций встречается наиболее часто;
• трансверсии -приводят к замене пурина пиримидином, и наоборот (сложная замена), т. е. вместо пары А–Т появляется пара Т–А или Г–Ц.
Возможны несколько генетических последствий точковых мутаций: 1) сохранение смысла кодона из-за вырожденности генетического кода (синонимическая замена нуклеотида); 2) изменение смысла кодона, приводящее к замене аминокислоты в соответствующем месте полипептидной цепи (миссенс-мутация; 3) образование бессмысленного кодона с преждевременной терминацией (нонсенс-мутация); 4) обратная замена стоп-кодона на смысловой кодон; 5) мутации сдвига рамки считывания.
Мутации со сдвигом рамки считывания представляют собой делеции или вставки одной или нескольких нуклеотидных пар. Большая часть изученных мутаций, вызывающих сдвиг рамки, обнаружена в последовательностях, состоящих из одинаковых нуклеотидов. Как следствие – изменяется последовательность аминокислот в белке мутантного штамма. Ревертанты в данном случае получить трудно.
Мутации, затрагивающие множество пар нуклеотидов, называют хромосомными. Они делятся на дупликации, делеции, инсерции, инверсии, транслокации. Дупликации – возникновение в данной нуклеотидной последовательности одного или, чаще, нескольких повторов. Делеции – утрата двух или нескольких пар оснований. Инверсии – изменение порядка нуклеотидов в ДНК на обратный по отношению к ориентации в штаммах дикого типа, они возникают обычно в результате рекомбинации с переворотом (flip-flop). Транслокации – перенос фрагмента ДНК в новое положение.
В настоящее время расшифрованы механизмы действия некоторых мутагенов.
Азотистая кислота (HNO 2) дезаминирует аденин, гуанин или цитозин, что приводит к ошибкам при репликации ДНК. Таким образом, происходит простая замена оснований, или транзиция. Например, если HNO2 взаимодействует с цитозином, то он дезаминируется с образованием урацила и при репликации образуется пара с аденином (вместо гуанина), что приводит к мутации ГЦ–АТ (транзиция).
Гидроксиламин (NH 2 OH) вступает в реакцию главным образом с цитозином и изменяет его так, что он при репликации ДНК предпочтительно спаривается с аденином вместо гуанина и происходит замена ЦГ–АТ (транзиция).
Аналоги азотистых оснований очень сходны по строению с нормальными пуриновыми и пиримидиновыми азотистыми основаниями и, поглощаясь клетками, способны включаться в ДНК. В молекуле ДНК они могут находиться в двух таутомерных формах – обычной кето-, или аминоформе, и реже встречающейся – енольной, или иминоформе. Переход в другую таутомерную форму может привести к неправильному образованию пар во время репликации ДНК. Часто для выделения мутантов используют 5-бромурацил (сходен с тимином) и 2-аминопурин (сходен с аденином).
Алкилирующие агенты – нитрозогуанидин, нитрозометилмочевина, этилэтансульфонат, этилметансульфонат, сернистый иприт и другие – принадлежат к наиболее эффективным мутагенам. Они алкилируют в области репликативной вилки преимущественно пуриновые основания, в первую очередь гуанин, вызывая его спаривание с тимином вместо цитозина. В результате этого возникают главным образом транзиции типа ГЦ–АT.
Молекулы акридиновых красителей (например, акридиновый оранжевый, акрифлавин, трипофлавин) внедряются между соседними азотистыми основаниями в цепи ДНК и увеличивают расстояние между ними. Такое изменение пространства при репликации ДНК может вызывать ошибки двух типов – утрату нуклеотида или включение дополнительной пары нуклеотидов. Мутации этого типа приводят к очень серьезным последствиям, так как при этом нарушается порядок считывания кодонов: начиная с места выпадения или вставки нуклеотида, информация считывается в «неправильных» триплетах, что приводит к формированию мутаций со сдвигом рамки считывания.
УФ-лучи действуют на тиминовые основания, следствием чего является образование димеров тимина в ДНК. Такие димеры служат источником возникновения ошибок при репликации ДНК. УФ-лучи вызывают мутации типа транзиций, трансверсий или делеций.
4. Мобильные генетические элементы бактерий
В качестве мутагенных факторов биологической природы рассматривают мобильные (= мигрирующие) генетические элементы бактерий – дискретные сегменты ДНК, способные к самостоятельному перемещению из одного участка в другой в пределах репликона, а также к перемещению из одного репликона (хромосомного, плазмидного или фагового) в другой. К таким элементам относятся: простые вставочные последовательности (IS-элементы), транспозоны (Tn-элементы) и фагитранспозоны (Mu, Д3112 и др.). Интеграция их в репликоны происходит независимо от системы общей рекомбинации клеток, которая требует обязательной гомологии у рекомбинирующих структур.
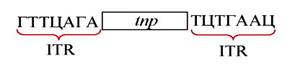
IS-элементы представляют собой линейные фрагменты двухцепочечной ДНК длиной от 200 до 2000 п. н. Они содержат только гены tnp, кодирующие синтез фермента транспозазы, необходимого для их миграции (транспозиции). По концам IS-элементов расположены инвертированные терминальные повторы (ITR). У разных IS-элементов длина концевых повторов ITR варьирует от 8 до 40 п. н. Инвертированные повторы также принимают участие и важны для транспозиции. Схематично строение IS- элемента можно изобразить следующим образом:
|
Различают несколько типов IS-элементов: IS1, IS2, IS3, IS4 и др. Они отличаются друг от друга по длине и структурой концевых повторов.
IS-элементы являются нормальными компонентами бактериальных хромосом и плазмид. В разных репликонах может содержаться различное, и часто множественное, число копий IS-элементов. IS-элементы могут перемещаться из одного участка генома в другой, например, из бактериальной хромосомы в плазмиду или от плазмиды к плазмиде. Также они могут встраиваться в пределах одного гена и инактивировать его или изменять его регуляцию.
Транспозоны – сложные мигрирующие элементы. Обозначаются как Tn 1, Tn 2,… Tn100, Tn 1002 и т.д. От IS-элементов они отличаются тем, что кроме генов, ответственных за транспозицию, содержат структурные гены, которые отвечают за проявление какого-либо фенотипа. Транспозоны могут контролировать резистентность к антибиотикам и ионам тяжелых металлов, способность к катаболизму лактозы, раффинозы, деградации толуола, синтезу энтеротоксинов и т. п., поэтому их легче обнаружить, чем IS-элементы. Длина транспозонов свыше 2000 п. н. Как и IS-элементы, транспозоны имеют инвентированные концевые повторы (ITR), которыми часто служат IS-элементы. Транспозоны различают не только по строению и составу, но и по степени специфичности при выборе мест интегрирования в репликоны. Однако следует отметить, что специфичность транспозиции одного и того же транспозона для разных видов бактерий и репликонов может быть различной.
Частота миграции транспозонов и IS-элементов происходит с вероятностью 10–4–10–7 на одно деление бактериальной клетки. Она может зависеть от характера донорного и реципиентного репликонов, а также от генома клетки-хозяина. Кроме того, на перемещение транспозонов могут влиять факторы внешней среды (температура, УФ-лучи, химические соединения и др.). Механизмы перемещения транспозонов окончательно не изучены.
Бактериофаг Mu относится к умеренным бактериофагам. Характерной его особенностью является мутагенность, что отражено в названии Mu (mu tator). Этот бактериофаг был впервые обнаружен у бактерий E. coli, но он размножается также на клетках Shigella, Klebsiella, Pseudomonas, Citrobacter, Salmonella и др. Он причисляется к мобильным генетическим элементам, так как во многих отношениях сходен с IS-элементами и транспозонами и отличается, по существу, только тем, что может формировать вирусные частицы. Сходство с IS-элементами и транспозонами в первую очередь выражается в том, что геном фага Mu (линейная двуспиральная ДНК – 38 т. п. н.) также имеет на концах инвертированные повторы, но только всего из двух нуклеотидных пар:
5′ ТГ ……………… ЦА 3′ОН
ДНК-Mu связана с геномом хозяина на протяжении всех случаев репликации и транспозиции как во время литического цикла после индукции профага, так и при заражении клеток извне. В этом отношении фаг Mu не отличается от других мигрирующих генетических элементов бактерий, но принципиально отличается от остальных умеренных фагов, которые вырезаются при индукции и размножаются вне связи с хозяйской хромосомой. После заражения клетки вирионами фага Mu его геном, прежде всего, интегрируется, и затем только в таком виде реплицируется, тогда как другие умеренные фаги в состоянии вегетативного фага могут размножаться без предварительной интеграции.
Значение мигрирующих генетических элементов:
1) Способны индуцировать образование мутаций. Интеграция IS-элементов и транспозонов также может привести и к прямо противоположному эффекту – экспрессии соседнего «молчащего» гена.
2) Участвуют в слиянии и диссоциации репликонов (например, в объединении трансмиссивных и нетрансмиссивных плазмид, в интеграции плазмид в хромосому и т. д.).
3) Наряду с плазмидами и фагами могут обеспечивать перенос генов между различными видами бактерий, иногда весьма отдаленными, и, следовательно, играют важную роль в эволюции микроорганизмов.
4) Используются в генной инженерии in vivo и in vitro.
5) Существенно ускоряют разработку частной генетики бактерий, которые имеют промышленное значение.
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 5031; Нарушение авторских прав?; Мы поможем в написании вашей работы!