КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Лекция № 11-12: Генетика пола. Скрепленное с полом наследование. Влияние внешней среды на генотип. Предупреждение наследственных заболеваний человека
|
|
|
|
Пол — это совокупность признаков и свойств организма, определяющих его участие в размножении. Пол особи может определяться:
а) до оплодотворения яйцеклетки сперматозоидом (прогамное определение пола)
б) в момент оплодотворения (сингамное определение пола);
в) после оплодотворения (эпигамное определение пола).
До оплодотворения пол определяется у некоторых организмов в результате разделения яйцеклеток на быстро и медленно растущие. Первые (более крупные) после слияния с мужской гаметой дают самок, а вторые (мелкие) — самцов. У коловраток, способных размножаться помимо обычного полового размножения с оплодотворением, партеногенетически, часть партеногенетических яйцеклеток во время развития лишается половины хромосом. Из таких яиц развиваются самцы, а остальная часть дает начало самкам.
У морского кольчатого червя бонеллия определение пола происходит в процессе онтогенеза: если личинка садится на дно, из нее развивается самка, а если прикрепляется к хоботку взрослой самки, то самец.
У подавляющего же большинства эукариот пол закладывается в момент оплодотворения и определяется генотипически хромосомным набором, который зигота получает от родителей. Клетки мужских и женских особей животных организмов различаются по паре хромосом. Эту пару называют половыми хромосомами (гетеросомами) в противоположность остальным — аутосомам. Половые хромосомы принято обозначать как Х- и Y-хромосомы. В зависимости от их сочетания у  и
и  организмов различают 5 типов хромосомного определения пола:
организмов различают 5 типов хромосомного определения пола:
1) XX, ХО (O обозначает отсутствие хромосом) встречается у видов Protenor (насекомые);
2) XX, XY — он характерен, например, для дрозофилы, млекопитающих (в том числе и для человека);
3) XY, XX — этот тип определения пола характерен для бабочек, птиц, рептилий;
4) ХО, XX — наблюдается у тли;
5) гапло-диплоидный тип ( 2n, n) встречается, например, у пчел: самцы развиваются из неоплодотворенных гаплоидных яйцеклеток, самки — из оплодотворенных диплоидных.
Конкретные механизмы, связывающие развитие мужского или женского пола с определенным сочетанием половых хромосом у разных организмов различен. У человека, например, пол определяется наличием Y-xpoмосомы: в ней есть ТДФ-ген, он кодирует тестикул - детерминирующий фактор, который определяет развитие мужского пола.
У дрозофилы же в Y-хромосоме находится ген фертильности, ответственный за плодовитость самца, а пол определяется балансом числа Х-хромосом и числа наборов аутосом (обычный диплоидный организм содержит, соответственно, два набора аутосом). В Х-хромосомах расположены гены, определяющие развитие по пути самки, а в аутосомах — по пути самца.
Если отношение количества Х-хромосом к количеству наборов аутосом равно 0,5, то развивается самец, а если — 1, то самка.
Помимо нормальных самцов и самок иногда появляются интерсексы — особи, по своим половым признакам занимающие промежуточное положение между мужским и женским полом (не путать с гермафродитами!). Это может быть вызвано как анеуплоидией по половым хромосомам в гаметах, так и различными нарушениями (например, гормональными) в процессе дифференцировки пола.
У большинства организмов среди потомков получается 50 % самцов и 50 % самок, так как в норме расщепление по полу подчиняется законам обычного моногибридного скрещивания между гомо- и гетерозиготой, для гибридов которого характерно расщепление в соотношении 1:1.
Наследование и пол
В половых хромосомах, помимо генов, определяющих развитие пола, локализуются «обычные» фенотипические гены. Особенности их наследования определяются тем, что они составляют группу сцепления гетеросом. Явление сцепленного с полом наследования было открыто Т. X. Морганом, который обнаружил, что наследование окраски глаз у дрозофилы находится во взаимосвязи с полом родителей — результаты прямого и обратного скрещивания были неодинаковы. Проведя ряд экспериментов, ученый пришел к выводу, что в Y-xpoмосоме самца не содержится участка, кодирующего окраску глаз.
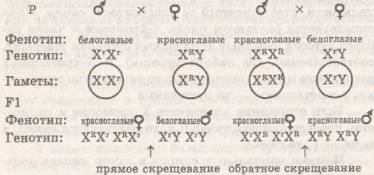
У человека сцепленными с полом являются такие аномалии, как дальтонизм и гемофилия. Поскольку рецессивные гены этих заболеваний локализованы в Х-хромосомах, ими чаще болеют мужчины; женщины же обычно гетерозиготны и по этой причине не болеют.
Если ген локализован в Y-хромосоме, ему в клетке нет гомологичной аллели, такой организм называют гемизиготой.
Некоторые гены могут находиться и не в половых хромосомах, однако их проявление будет зависеть от пола особи: у одного пола признак проявится, у другого — нет. Такие признаки называют признаками, ограниченными полом. К ним относятся, например, наличие рогов у оленей (самцы рогаты, а самки безроги) или яйценоскость птиц, которая проявляется только у самок. Обычно проявление признака, ограниченного полом, зависит от гормонального статуса организма, в первую очередь, от соотношения половых гормонов.
Воздействие внешней среды на генотип
Мутационная изменчивость проявляется в фенотипе, и по сути дела лишь по наличию качественно новых признаков и свойств организма можно предполагать ее возникновение. Изменения фенотипа вызываются нарушением наследственных структур, которое обусловливается влиянием различных факторов внешней среды.
Иными словами, внешняя среда, воздействуя на генотип, вызывает его структурные изменения, приводящие к формированию новых признаков и свойств организма.
В связи с этим исследование мутаций должно вестись с разных позиций: с точки зрения характера изменений в генотипе, локализации их в различных клетках и тканях, фенотипического выражения и эволюционной роли мутаций, а также с точки зрения природы причинного фактора. Существует много классификаций мутаций. Наиболее удобна классификация, разработанная польским исследователем С. Мушинским в 1972 г. Пользуясь этой классификацией, можно получить представление о мутации на молекулярном и цитологическом уровнях, о ее локализации в клетках и тканях, о фенотипическом проявлении и судьбе в популяции.
Молекулярный механизм мутаций. Мутации, связанные с изменением структуры молекулы ДНК, называются генными. Они представляют собой выпадение или вставку одного или нескольких азотистых оснований либо то и другое одновременно, а также замену азотистых оснований. Последние описаны Э. Фризом (1963). Он различал два типа замен: транзиции и трансверзии.
При транзиции одно пуриновое или пиримидиновое основание заменяется соответственно другим пуриновым или пиримидиновым основанием.
Трансверзии встречаются чаще транзиций.
Все генные мутации приводят к изменению смысла кодона и к нарушению считывания информации в цепи ДНК. Различают следующие типы таких изменений.
Миссенс-мутации, т. е. мутации, изменяющие смысл кодона, вследствие чего в белковую молекулу в момент ее синтеза вставляется другая аминокислота.
Нонсенс-мутации — образование бессмысленных кодонов, не кодирующих никакой аминокислоты (УАА — охра-мутация; УАГ — амбер, или янтарная мутация; УГА — опал-мутация). Такие мутации приводят к обрыву чтения генетического текста и прекращению синтеза молекулы белка.
Миссенс и нонсенс-мутации обычно происходят при замене азотистых оснований. К изменению смысла кодонов приводят и выпадения или вставки азотистых оснований. Все эти мутации возникают спонтанно и могут быть вызваны любыми мутагенными факторами среды.
Мутации сдвига чтения наблюдаются при выпадении или вставке нуклеотидов в цепи ДНК и вызывают смещение чтения генетического кода. При этом рано или поздно образуются бессмысленные кодоны, на которых чтение прерывается.
Хромосомные мутации. В клетке под обычным световым микроскопом можно рассмотреть хромосомные мутации, или аберрации. Они являются более грубыми нарушениями наследственных структур, чем генные мутации, и касаются структуры и количества хромосом в клеточном наборе.
Структурные хромосомные мутации связаны с нарушением целостности структуры хромосомы, групп сцепления генов, с процессом ее фрагментации. Эти мутации бывают двух типов: внутрихромосомные, изменяющие порядок расположения генов в хромосоме, и межхромосомные, заключающиеся во взаимном обмене фрагментов хромосом. Обычно для формирования структурной мутации требуются два и более разрыва хромосомы, в некоторых случаях достаточно одного. Различают хромосомные и хроматидные аберрации. Если разрыв затрагивает одну хроматиду, перестройка называется хроматидной, но после репликации она может стать хромосомной.
К внутрихромосомным перестройкам относятся делеции, дупликации, инверсии и инсерции.
Делеция — это потеря (нехватка) среднего участка хромосомы вследствие ее разрыва в двух точках. Если происходит отрыв дистального, концевого, фрагмента, нехватка называется дефишенси. При потере хроматидами двух таких районов последние соединяются в местах повреждения и образуют кольцевую хромосому. Дефишенси встречаются редко, так как после потери дистального района хромосома неспособна к дальнейшему существованию. Судьба делеции и дефишенси зависит также от того, теряется ли центромерный район. Без центромера хромосома в процессе деления элиминируется. При конъюгации хромосом в мейозе или при соматической конъюгации в полигенных хромосомах двукрылых нормальная хромосома образует над участком делеции аберрантной хромосомы петлю, поскольку ее фрагмент не имеет в данном случае гомолога.
Нехватки обычно понижают жизнеспособность и плодовитость особи. Часто клетка с делецией гибнет на ранних стадиях развития. Мелкие нехватки могут сохраняться в гомозиготном состоянии, проявляя фенотипическое тождество с генной мутацией. Вместе с тем в случае делеции невозможен возврат гена к исходному состоянию, т. е. обратная мутация исключается.
Дупликация — удвоение фрагмента хромосомы — процесс, противоположный делеции. При конъюгации дуплицированная хромосома также делает над нормальной хромосомой петлю, которая в отличие от делеции несет дуплицированные гены. Примером дупликации является усиление признака Ваг (полосковидные глаза) у дрозофилы при увеличении числа генов, контролирующих его.
Явление дупликации генов сравнительно часто встречается в природе, и ему приписывается определенная эволюционная роль.
Нередко при разрывах хромосомы ее фрагменты не утрачиваются, а, повернувшись на 180°, встраиваются в то же место. Такая мутация называется инверсией. Интересным примером инверсий служат различия хромосомных наборов в семействе кошачьих. Все его представители имеют 36 хромосом, но кариотипы разных видов отличаются наличием инверсии в различных хромосомах.
Инверсии приводят к изменению ряда морфологических и физиологических признаков организма, могут явиться фактором биологической изоляции популяции. Процесс конъюгации хромосомы с инверсией затрудняется, что вызывает нарушения мейоза и тормозит кроссинговер. Поэтому инверсии часто служат «запирателями» кроссинговера.
Инсерция — перемещение фрагментов хромосомы по ее длине, замена локализации генов. Такая аберрация часто сопровождается эффектом положения генов.
Межхромосомные перестройки представляют собой еще один тип мутаций — транслокации. Они возникают при одновременном разрыве в разных хромосомах, которые затем обмениваются фрагментами.
При взаимном обмене примерно равными участками двух гомологичных хромосом происходит реципрокная транслокация.
При одностороннем переносе фрагмента одной хромосомы на другую возникает нереципрокная транслокация. Грубые транслокации могут привести к резкому снижению жизнеспособности клетки и организма в целом. Однако нередко встречаются организмы, несущие транслокации, но сохраняющие жизнеспособность. Наличие в природных популяциях особей с инверсией или транслокацией препятствует получению нормального потомства при скрещивании их с нормальными особями и является генетическим фактором изоляции популяции.
Профилактика наследственных заболеваний. Евгеника – наука об улучшении признаков и свойств, наследуемых человеком, и недопустимости браков, от которых рождается неполноценное потомство. Этот термин был введен в 1883 году английским ученым Ф.Гальтоном. Евгеника начала развиваться в Европе и США. В некоторых штатах Америкибыли введены законы евгеники. Использование современных геномных технологий для профилактики наследственных заболеваний привело, с одной стороны, к повышению эффективности медико-генетической помощи, а с другой - к необходимости решения ряда биоэтических проблем. Это связано с тем, что в генетическом исследовании бывает задействовано много сторон: исследователь, врач-клиницист, испытуемый, донор, реципиент, пациент и члены его семьи. Косвенно в этих исследованиях могут участвовать и некоторые социальные службы: образования, трудоустройства, страхования (жизни, здоровья, имущества) и прочие, где люди вступают в отношения друг с другом в связи с проведением генетических исследований или приложением их результатов в своей сфере практической жизни. При этом люди могут по-разному работать с образцами биоматериала: осуществлять их забор и изучение, технологически трансформировать, передавать их другим лицам (исследователям, врачам, кому-либо еще), вводить в организм реципиента образцы, содержащие генетическую информацию. Причем информацию они тоже могут использовать по-разному: хранить, передавать, распространять, уничтожать. Но к данной генетической информации причастны и те лица, которые являются испытуемыми, пациентами, членами их семей и т.д. При этом у этики генетики есть одно отличие от многих других разделов биомедицинской этики, а именно: не только сам испытуемый, но и его прямые потомки в нескольких поколениях могут оказаться объектами воздействия измененной генетической информации. В основе решения проблем, возникающих при получении и использовании генетической информации обследуемого индивида должны лежать следующие принципы:
1. Признание автономии личности. Важно, что все виды процедур должны осуществляться с информированного согласия объекта, которое означает, что человек вступает в контакт с генетиком, врачом или иным исследователем добровольно. При этом профессионал обязан обеспечить его в доступной форме адекватной информацией, чтобы она была и необходима, и достаточна, и понятна, и помогала бы автономной личности принимать самостоятельное решение о том, на что (на какие процедуры) она (личность) согласна пойти, а на что - не согласна. Приоритеты в этическом решении о проведении генетических процедур убывают в гаком направлении: индивид, семья и кровные родственники, общество.
2. Гиппократовское «не вреди» и «сотвори благо», направленное на сохранение конфиденциальности информации. Это правило проходит через все документы по биоэтической регламентации любой медицинской деятельности. Согласно этому правилу информация о генетическом статусе человека может быть сообщена только ему, его опекунам или иным легальным представителям и пользующим его врачам. Недопустима передача профессионалом какой-либо информации без санкции тестируемого или его законных опекунов любой третьей стороне (органам образования, трудоустройства, страхования, социальным службам и т.д.), так как это может повлечь за собой дискриминацию такого лица на основании сведений о его генетическом статусе, что равносильно своего рода «генетическому шовинизму»,
3. Справедливость, которая подразумевает равные возможности генетического тестирования за счет общественных фондов для всех членов общества.
Вопросы для закрепления темы:
1. Каким способом можно определить пол человека?
2. Что такое аутосомы, половые хромосомы?
3. Какие перестройки относятся к внутрихромосомным?
4. Какие генные мутации вы знаете? Как они подразделяются?
5. Какими значками обозначаются хромосомы?
6. Какое влияние оказывает внешняя среда на изменение генотипа?
7. Как предотвратить передачу наследственных заболеваний?
8. Возможно ли заболевание гемофилией у женщин?
9. Какому поколению передается крисс-кросс наследование?
10. Как вы понимаете гомогаметность и гетерогаметность?
11. Что изучает наука евгеника?
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 1496; Нарушение авторских прав?; Мы поможем в написании вашей работы!