КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Субстраты дыхания и дыхательный коэффициент
|
|
|
|
В качестве основного субстрата дыхания растения используют углеводы, причем в первую очередь окисляются свободные сахара. При их недостатке могут быть использованы полисахариды, белки, жиры после их гидролиза. Поли- и дисахариды гидролизуются до моносахаридов, белки - до аминокислот, жиры - до глицерина и жирных кислот.
Использование жиров начинается с их гидролитического расщепления липахой до глицерина и жирных кислот, что происходит в сферосомах. Благодаря фосфорилированию и последующему окислению глицерин превращается в фосфотриозу – ФГА, который включается в основной путь обмена углеводов.
Жирные кислоты окисляются по механизму β-окисления, в результате которого от жирной кислоты последовательно отщепляются двууглеродные ацетильные остатки в форме ацетил-СоА. Данный процесс происходит в глиоксисомах, где, кроме того локализованы ферменты глиоксилатного цикла. Ацетил-СоА включается в реакции глиоксилатного цикла, конечный продукт которого – сукцинат – покидает глиоксисому и в митохондриях участвует в цикле Кребса (рис.). Синтезированный в ЦТК малат в цитоплазме при участии малатдегидрогеназы превращается в оксалоацетат, который с помощью ФЕП-карбоксилазы дает ФЕП. ФГА и ФЕП служат исходным материалом для синтеза глюкозы (а также фруктозы и сахарозы) в в обращенных реакциях гликолиза. Процесс образования глюкозы из неуглеводных предшественников получил название глюконеогенеза.. Экспериментально доказано, что по мере прорастания в семенах снижается содержание жиров и увеличивается – сахаров.
Запасные белки используются для дыхания в результате гидролиза до аминокислот и последуешего окисления до ацетил-СоА или кетокислот, которые затем поступают в цикл Кребса (рис.)
Полное окисление рассмотренных субстратов осуществляется до углекислого газа и воды с освобождением энергии окисляемых веществ.
Отношение количества молей СО2 выделяемого при дыхании к количеству молей поглощенного О2 называется дыхательным коэффициентом (ДК). Для гексоз он равен единице:/
С6О12О6 + 6О2 → 6СО2 + 6Н2О; ДК= 6СО2/6О2=1
Количество кислорода, необходимое для окисления субстрата, находится в обратной зависимости от содержания его в молекуле субстрата. Поэтому если субстратом дыхания являютяс более бедные кислородом (по сравнению с углеводами)жирные кислоты, то ДК будет меньше елиницы:
С18Н36О2 + 26О2 → 18СО2 + 18Н2О; ДК=18 СО2/26 О2=0,69
На величину ДК влияют и другие факторы, например, недостаток кислорода (при затоплении корней и др.) усиливается брожение и ДК возрастает; если в результате недоокисления продуктов в тканях накапливаются органические кислоты, а количество углекислого газа снижается, ДК падает.
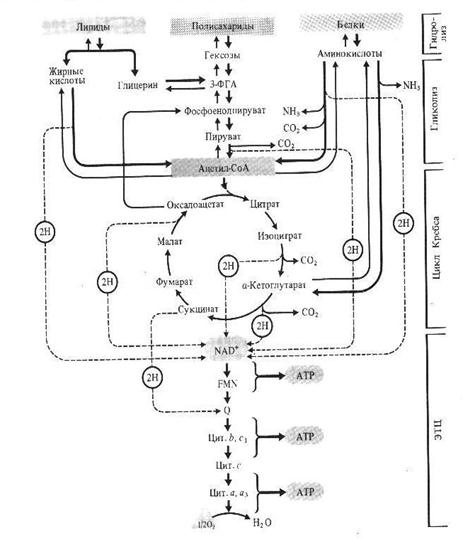
Рис. Использование полисахаридов, белков и жиров в качестве дыхательных субстратов.
- Зависимость дыхания от факторов внешней среды
1. Концентрация кислорода
Процесс дыхания связан с непрерывным потреблением кислорода. Но окислительные превращения субстратов включают аэробные и анаэробные процессы (гликолиз, брожение). Снижение парциального давления кислорода с 21% до 5% интенсивность дыхания тканей меняется незначительно.
Впервые влияние кислорода на величину расходования дыхательных субстратов обнаружил Л.Пастер. В его опытах с дрожжами в присутствии кислорода снижались распад глюкозы и интенсивность брожения, но одновременно наблюдался интенсивный рост биомассы. Торможение распада сахаров и более эффективное их использование в присутствии кислорода получило название «эффекта Пастера».. Это объясняется тем, что при высоком парциальном давлении кислорода весь пул ADP и Р расходуется на синтез АТР. В результате происходит торможение гликолиза из-за снижения количества ADP и Р, необходимых для субстратного фосфорилирования и высокое содержание АТР угнетает некоторые гликолитичесие ферменты (фосфофруктокиназу). В итоге снижается интенсивность гликолиза и активируются синтетичесие кроцессы (глюконеогенез)
Важным фактором, определяющим интенсивность дыхания клетки, является концентрация ADP. Зависимость скорости потребления кислорода от концентрации ADP, называется дыхательным контролем, или акцепторным контролем дыхания. Соотношение суммы концентраций АТР и 1/2ADP к сумме концентраций АТР, ADP, AMP называют энергетическим зарядом.
Избыток кислорода в тканях растений может возникать лишь локально. В атмосфере чистого кислорода дыхание растений снижается, а затем растение погибает. Это связано с усилением в клетках свободнорадикальных реакций, окислением липидов мембран, и, как следствие – нарушением всех обменных процессов.
2. Концентрация углекислого газа
Повышение концентрации СО2 приводит к снижению интенсивности дыхания, т.к. тормозятся реакции декарбоксилирования и активность сукцинатдегидрогеназы. При наблюдается закисление тканей – ацидоз.
3. Температура
Дыхание, как ферментативный процесс, зависит от температуры. В определенных температурных границах эта зависимость подчиняется правилу Вант-Гоффа (скорость химических реакций удваивается при повышении температуры на 10оС). Для дыхания каждого вида растений и его органов существуют определенные минимальные, оптимальные и максимальные температуры.
4. Водный режим
В листьях проростков при быстрой потере воды в начале отмечается усиление дыхания. При постепенном снижении обводненности этого не происходит. Длительный водный дефицит приводит к снижению дыхания. Особенно отчетливо влияние воды прослеживается при изучении дыхания семян. При повышении влажности семян до 14-15% дыхание возрастает в 3-4 раза, до 30-35% - в тысячи раз. При этом важную роль играет температура.
5. Минеральное питание
Добавление раствора солей, в воду, где выращивались проростки, обычно усиливает дыхание корней. Этот эффект получил название «солевого дыхания». В тканях других органов этот эффект удается получить не всегда
- Повреждения и механические воздействия
Механические воздействия вызывают кратковременные усиления поглощения кислорода по трем причинам: 1) из-за быстрого окисления фенольных и других соединений, которые выходят из вакуолей поврежденных клеток и становятся доступными для соответствующих оксидаз; 2) в связи с увеличением количества субстрата для дыхания; 3) вследствие активации процессов восстановления мембранного потенциала и поврежденных клеточных структур.
|
|
|
Дата добавления: 2013-12-13; Просмотров: 5618; Нарушение авторских прав?; Мы поможем в написании вашей работы!