КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Пресмыкающиеся
|
|
|
|
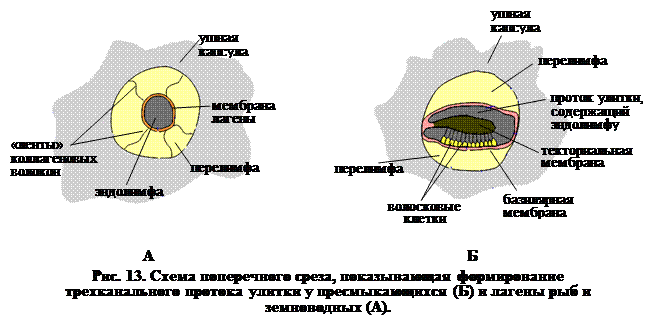
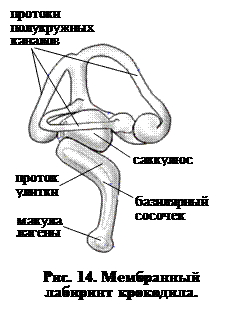
Именно у пресмыкающихся происходит закладка настоящей улитки, которая приобретает такое же значение как у птиц и млекопитающих. У аллигаторов, например, лагена (вырост саккулюса) присоединяется к стенке окружающей ее полости уха, так что образуются три различных канала (рис. 13). Параллельно увеличивается базилярный сосочек (рис. 14), который состоит из полоски волосковых клеток, вырастающей из основной мембраны протока улитки, а начало улитки покрыто непрерывной желатинозной мембраной. Эта текториальная мембрана играет важную роль в восприятии вибраций.
 Зачатки этого обнаруживаются и у земноводных, но у рептилий он развит значительно лучше, особенно у крокодилов и аллигаторов. Наиболее важную роль играет у птиц и млекопитающих.
Зачатки этого обнаруживаются и у земноводных, но у рептилий он развит значительно лучше, особенно у крокодилов и аллигаторов. Наиболее важную роль играет у птиц и млекопитающих.
Барабанная перепонка часто расположена глубже поверхности тела, так что возникает зачаток наружного слухового прохода, и, как у бесхвостых амфибий, слуховой стержень простирается через камеру среднего уха, проводя вибрации в мембранный лабиринт внутреннего уха.
Змеи являются исключением из всех пресмыкающихся, поскольку не имеют ни наружного слухового прохода, ни камеры среднего уха. У них однако имеется слуховой стержень, который присоединяется к квадратной кости челюстного сустава. Таким образом, змеи нечувствительны к колебаниям воздуха, но великолепно ощущают колебания почвы.
Хотя анатомические данные демонстрируют некоторый эволюционный прогресс пресмыкающихся по отношению к земноводным, мало что свидетельствует о лучшем качестве их слуха. Черепахи очень чувствительны к звукам в диапазоне 80-130 Гц, аллигаторы реагируют на частоты выше 1000 Гц, ящерицы ощущают колебания частотой до 10 кГц, но нет оснований предполагать, что они способны хорошо различать частоты.
3.2.4.  Птицы
Птицы
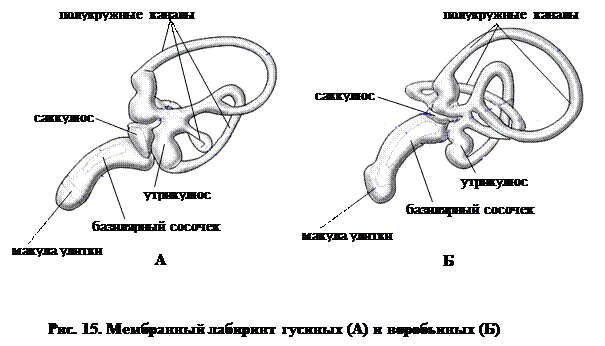
Анатомия органа слуха у птиц развивалась сходным образом с таковым у млекопитающих (рис. 15). Улитка значительно удлиняется и приобретает трехканальную структуру, которая впервые возникает, как мы видели выше, у пресмыкающихся:
ü верхний канал называется вестибулярной лестницей,
ü средний – средней лестницей,
ü нижний – барабанной лестницей.
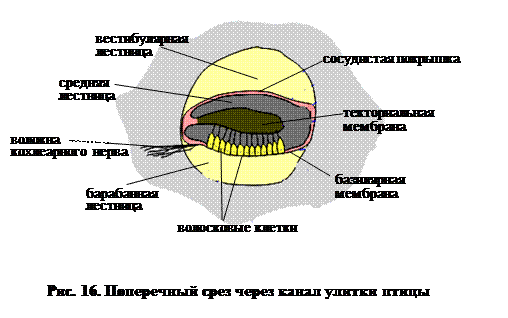
Поперечный срез (рис. 16) показывает, что базилярный сосочек увеличивается и приобретает структуру, называемую кортиевым органом. Ширина базилярной мембраны увеличивается по направлению к ее дистальному концу, что предполагает детектирование низкочастотных тонов именно в этом месте. За дальним концом улитки сохраняется макула лагены, которая также участвует в восприятии тонов низкой частоты.
Таким образом, ухо у птиц состоит из трех отделов:
Ø наружного слухового прохода, ведущего к барабанной перепонке,
Ø среднегоуха, пересеченного слуховым стержнем – тремя неправильной формы косточками,
Ø внутреннего уха, содержащего мембранный лабиринт.
Большинство птиц, естественно, чрезвычайно чувствительны к звуку, который играет огромную роль в их социальном поведении. Песни птиц видоспецифичны, причем ясно, что птенцы выучивают ее локальный тип. Выяснилось, что некоторые из птиц, обитающих в пещерах, используют звуки и для эхолокации (венесуэльский гуахаро, азиатский стриж), однако нет никаких данных, которые бы свидетельствовали о том, что у птиц развита чувствительность к ультразвуковым колебаниям, которая характерна для летучих мышей. Наконец, чувствительность к направлению звука достигает очень высокого уровня у ночных хищников, таких как совы.
3.2.5. Млекопитающие.
У млекопитающих слуховая часть уха – улитка – достигает эволюционной вершины.
Самый слабый звук, который способен уловить человек, соответствует около 0,0001 дин/см2 или приблизительно 10-16 Вт/см2. Это соответствует изменению давления на барабанной перепонке в 20 мкПа. Диапазон частот, которые способен ощущать человек, простирается от 20 Гц до 20 кГц, хотя с возрастом верхняя граница чувствительности снижается. Тренированное ухо может различать частоты 1000 и 1002 Гц. Такая поразительная способность – следствие высокого развития улитки.
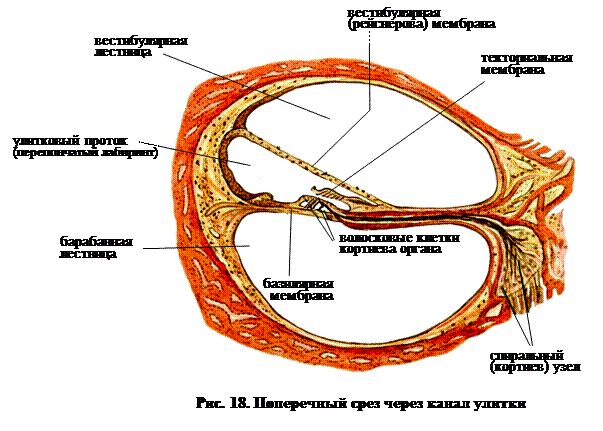
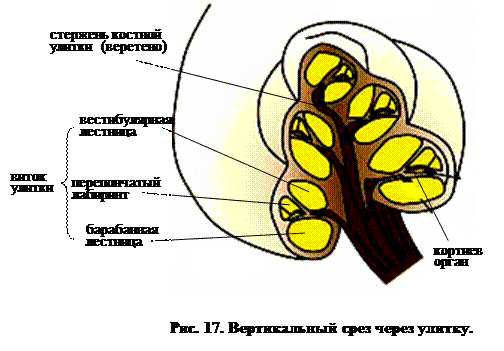 Как показывает рис. 9 Ж, у млекопитающих улитка уже не прямая или слабо изогнутая, как у птиц, а свернута в спираль, похожую по форме на раковину моллюска. Срез ее показан на рис. 17. Из рисунка видно, что три канала – вестибулярная, средняя и барабанная лестницы, с которыми мы впервые встречались у пресмыкающихся, присутствуют и здесь, а рис. 18 показывает, что базилярный сосочек развивается в сложный кортиев орган.
Как показывает рис. 9 Ж, у млекопитающих улитка уже не прямая или слабо изогнутая, как у птиц, а свернута в спираль, похожую по форме на раковину моллюска. Срез ее показан на рис. 17. Из рисунка видно, что три канала – вестибулярная, средняя и барабанная лестницы, с которыми мы впервые встречались у пресмыкающихся, присутствуют и здесь, а рис. 18 показывает, что базилярный сосочек развивается в сложный кортиев орган.
На рис. 19 показпно, что и среднее ухо (как и наружное) претерпело значительное развитие. У многих видов ушная раковина (не показана) велика и достаточно подвижна. Это играет значительную роль в определении направления на источник звука. У людей мышцы, двигающие ушную раковину в отличие от животных, сохраняются в основном в рудиментарной форме. Наружный слуховой проход сравнительно длинный и, для предупреждения попадания различных посторонних частиц, покрыт волосами и железами, секретирующими ушную серу.
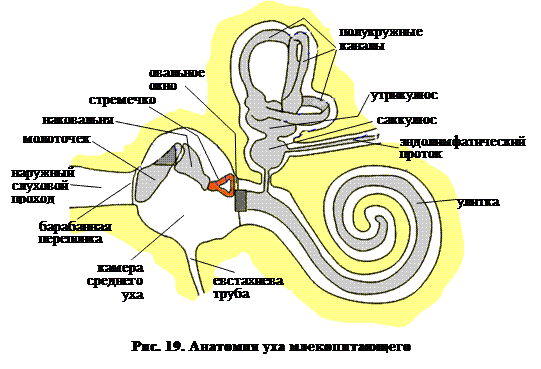
Барабанная перепонка представляет собой границу между наружным и средним ухом. В среднем ухе поддерживается атмосферное давление за счет наличия евстахиевой трубы, соединяющей полость среднего уха и открывающейся в глотку. Это важно, поскольку барабанная перепонка тонка, и резкое различие давления по обе стороны от нее может привести к ее разрыву. Вибрация барабанной перепонки, вызываемая звуком, передается через камеру среднего уха слуховыми косточками: молоточком, наковальней и стременем. Эти самые маленькие кости в организме человека прикреплены к стенке камеры среднего уха связками и мышцами. Мышца, идущая к молоточку, и мышца, идущая к стремечку, сокращаются одновременно в ответ на звуки высокой интенсивности и, таким образом защищают тонкие структуры внутреннего уха от повреждающего эффекта избыточной вибрации. Наконец, основание стремечка, овальная пластинка сочленяется с мембраной, закрывающей овальное окно круговой связкой. При развитии отосклероза эта часть стремени срастается с костным окружением овального окна, что ведет к глухоте.
Для упрощения рис. 20 показывает улитку, развернутую из спирали в прямую. Когда волна атмосферного давления, которую мы воспринимаем как звук, достигает барабанной перепонки, вибрация передается в овальное окно. Таким образом, мембрана, покрывающая окно, передает изменения давления перилимфе внутреннего уха (заполняющей вестибулярную и барабанную лестницы). Теперь рассмотрим рис. 18 и 20. Поскольку мембрана, покрывающая овальное окно смещается внутрь, давление в вестибулярной лестнице возрастает. Рейснерова мембрана сдвигается вниз, что увеличивает давление в средней лестнице до тех пор, пока оно не компенсируется смещением базилярной мембраны. Возникающее вследствие этого избыточное давление в барабанной лестнице компенсируется изгибанием наружу мембраны, покрывающей круглое окно.
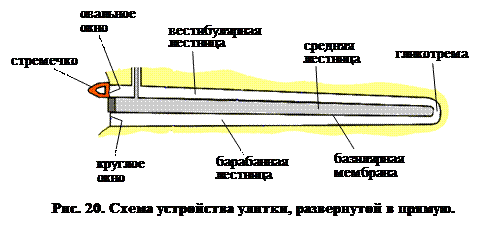
Как видно из рис. 18, в рассматриваемой системе присутствует еще одна – текториальная мембрана, опирающаяся на два ряда волосковых клеток. Кончики ресничек погружены в нее, точно также, как в описанном выше случае – в желатинозную купулу. В отличие от рейснеровой и основной, текториальная мембрана сравнительно свободно плавает в эндолимфе и следовательно изменения гидростатического давления влияют на нее в равной степени со всех сторон. Поэтому изменения давления, особенно быстрые изменения, не могут заставить эту мембрану двигаться вверх или вниз, синхронно с другими двумя.
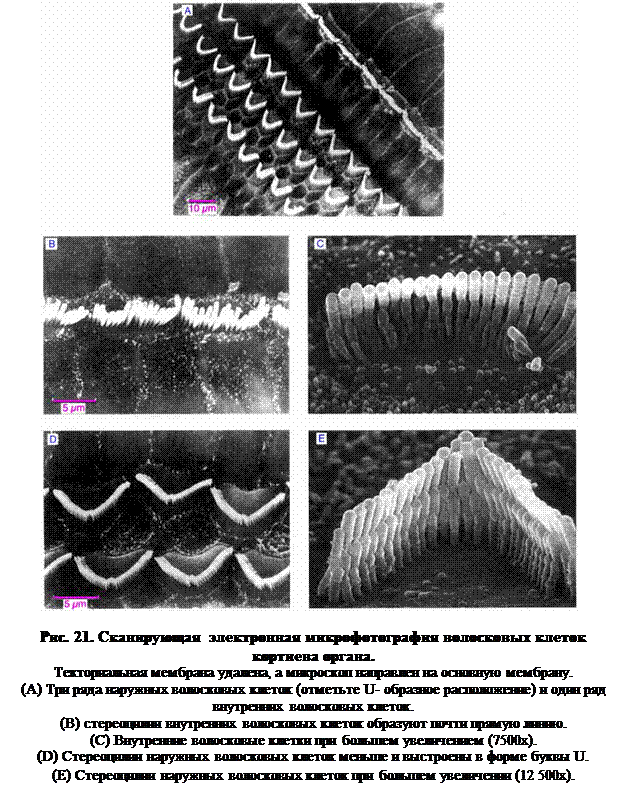
Теперь рассмотрим волосковые клетки. Среди них присутствует один ряд внутренних волосковых клеток, отделенных небольшим пространством от трех или четырех рядов наружных волосковых клеток. У этих клеток развивается от 50 до 100 стереоцилий, однако, в отличие от уже рассмотренных волосковых клеток, здесь нет киноцилий. На том месте, где можно было бы ожидать присутствия киноцилий, развивается крупная центриоль. Вид клетки снаружи показывает, что стереоцилии клеток располагаются таким образом, что принимают форму буквы «W» или «U» (рис. 21).
Кончики стереоцилии погружены в текториальную мембрану, а основания волосковых клеток присоединены к базилярной мембране, где они образуют синаптические контакты с дендритическими окончаниями кохлеарного нерва.
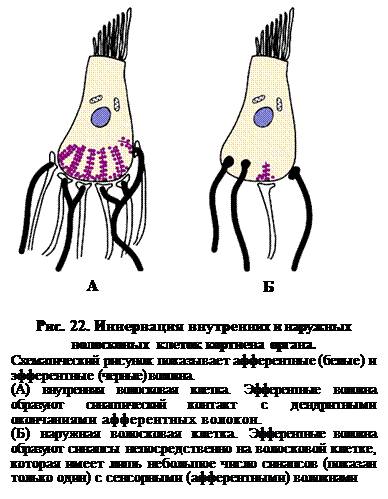
Внутренние волосковые клетки кортиева органа (рис. 22) содержат множество синаптических везикул (вероятно, содержащих глутамат), концентрирующихся вокруг электронно-плотного материала, который служит для организации везикул перед их выбросом. Синапсы имеют различные механизмы в пресинаптической зоне, обеспечивающие быстрое и эффективное выделение медиатора. Здесь концентрируются Са2+-каналы такого типа, которые способны быстро реагировать на малые изменения потенциала. Это обстоятельство обеспечивает и почти мгновенное прекращение выброса медиатора.
Между внутренними и наружными волосковыми клетками есть существенное различие. Тогда как внутренние волосковые клетки образуют контакты с приблизительно десятком афферентных волокон кохлеарного нерва, наружные имеют гораздо меньшую степень иннервации.
Около 90% афферентных волокон кохлеарного нерва являются продолжениями синаптических окончаний, образованных на ряде внутренних волосковых клеток.
Рис. 22 показывает, что основания волосковых клеток содержат множество синаптических пузырьков (везикул), а также «электронно-плотное тело» (синаптические ленты) и другие дополнительные структуры. Присутствуют также эфферентные волокна кохлеарного нерва. Эти волокна образуют синаптические контакты с дендритными окончаниями афферентных волокон. Из этого можно предположить, что имеет место контроль чувствительности с помощью системы обратных связей.
Наружные волосковые клетки, напротив, бедно иннервированные афферентными волокнами, получают мощную эфферентную иннервацию. Окончания эфферентных волокон – крупные, заполненные медиаторными везикулами. Кроме того, имеются данные о том, что наружные волосковые клетки обладают сократительной способностью. Вероятно, такая эфферентная иннервация также позволяет мозгу контролировать чувствительность улитки путем изменения расстояния между текториальной и базилярной мембранами.
Как известно, базилярная мембрана колеблется в ответ на поступающий в ухо звук, тогда как текториальная мембрана остается сравнительно неподвижной. Стереоцилии волосковых клеток подвергаются механической деформации, причем их реснички погружены в богатую К+ эндолимфу. Возникающая деполяризация может быть обнаружена с помощью микроэлектродных отведений. Они точно воспроизводят частоту поступающего звука. Это т.н. микрофонные потенциалы. Микрофонные деполяризации (рецепторные потенциалы) приводят к выделению медиаторных веществ на дендритные окончания афферентных волокон кохлеарного нерва.
Таким образом, очевидно, что в основе потрясающе сложного внутреннего уха млекопитающих лежат волосковые клетки; конечно, модифицированные, но в целом те же самые, которые обнаружены в каналах органа боковой линии наших водных предшественников.
Движущей силой в развитии улитки млекопитающих, была необходимость отличать разные частоты звука. В небольшой степени эта способность присутствует у рыб, земноводных и пресмыкающихся; у птиц и млекопитающих она претерпевает огромное развитие. Как уже было отмечено, частотный диапазон человеческого уха лежит между 20 Гц и 20 кГц (с некоторым снижением верхнего предела с возрастом). Кроме того, в пределах диапазона слышимости человек и другие млекопитающие обладают чрезвычайно высокой способностью к различению частот.
Для осуществления данной функции существуют два основных механизма:
1. Есть данные, что кохлеарные волокна могут быть фазо-синхронными звуковым частотам выше 500 Гц, но не реагируют на каждый частотный импульс. То есть предполагается, что в нижней части частотного спектра (ниже 5 кГц) группа волокон кохлеарного нерва объединяется для достижения частоты импульсов, совпадающей с тональной частотой, в каком-то слуховом центре мозга. По понятным причинам, такая идея именуется теорией залпа. Иными словами, в пучке нервных волокон кохлеарного нерва каждое волокно генерирует сигнал с определенной частотой таким образом, что, в результате, группа нервных волокон вместе дает импульсацию с частотой, совпадающей с таковой в каком-то слуховом центре мозга.
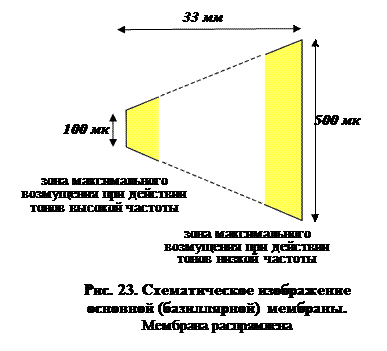 2. Основывается на наблюдении, что ширина базилярной мембраны увеличивается от круглого окна к геликотреме (или в случае птиц – к макуле улитки). Ширина базилярной мембраны человека, например, увеличивается с 100 до 500 мкм на расстоянии в 33 мм (рис. 23). Герман фон Гельмгольц еще в XIX веке предположил, что базилярную мембрану можно уподобить ряду настроенных камертонов (резонаторов). Тоны высокой частоты вызывают максимальные возмущения в области круглого окна, а низкой – у геликотремы. Точные исследования фон Бекеши в основном подтвердили гипотезу Гельмгольца. Обнаружено, что волны сложной формы двигаются вдоль всей базилярной мембраны, но место, где они достигают максимальной амплитуды, как и предположил Гельмгольц, связано с их частотой. Догадка Гельмгольца по очевидным причинам известна как теория места
2. Основывается на наблюдении, что ширина базилярной мембраны увеличивается от круглого окна к геликотреме (или в случае птиц – к макуле улитки). Ширина базилярной мембраны человека, например, увеличивается с 100 до 500 мкм на расстоянии в 33 мм (рис. 23). Герман фон Гельмгольц еще в XIX веке предположил, что базилярную мембрану можно уподобить ряду настроенных камертонов (резонаторов). Тоны высокой частоты вызывают максимальные возмущения в области круглого окна, а низкой – у геликотремы. Точные исследования фон Бекеши в основном подтвердили гипотезу Гельмгольца. Обнаружено, что волны сложной формы двигаются вдоль всей базилярной мембраны, но место, где они достигают максимальной амплитуды, как и предположил Гельмгольц, связано с их частотой. Догадка Гельмгольца по очевидным причинам известна как теория места  частотной дискриминации. Чтобы различить частоты, мозгу достаточно лишь «посмотреть», из какого места базилярной мембраны происходят волокна, в которых активность максимальна.
частотной дискриминации. Чтобы различить частоты, мозгу достаточно лишь «посмотреть», из какого места базилярной мембраны происходят волокна, в которых активность максимальна.
В последнее время теория места Гельмгольца была расширена и подкреплена обнаружением того факта, что сами по себе волосковые клетки настроены на определенную частоту звука. Такая настройка – результат молекулярной анатомии и цитологии клетки. Показано, что волосковые клетки и их стереоцилии различаются в размерах и гибкости в разных частях базилярной мембраны:
Ø вблизи от круглого окна стереоцилии малы и сравнительно жестки,
Ø в области геликотремы стереоцилии больше в размерах и гибки.
Это вместе со специфической популяцией ионных каналов определяет электрические характеристики данной клетки.
Каждая волосковая клетка настроена на максимальную реакцию в ответ на стимул определенной частоты. Есть и свидетельства того, что настройка клеток находится под контролем обратной связи от улитки. Сложная электромеханическая настройка волосковых клеток с модуляцией через обратные связи накладывается на механизм максимального возмущения в специфической части базилярной мембраны, связанной с частотой поступающего звука.
Многие млекопитающие чувствительны к звукам намного больших частот, чем способно различить человеческое ухо. Эксперименты по выработке условных рефлексов при воздействии на вибриссы или ушную раковину показали, что многие мелкие млекопитающие чувствительны к звукам частотой до 100 кГц. Слышимый для человека писк мышей и землероек находится в низкочастотной части спектра слышимости этих животных. Большая часть социальной коммуникации между этими млекопитающими реализуется за пределами слышимости для человека. У собак частотная характеристика также гораздо шире, чем у человека – их верхний предел находится в районе 35 кГц (что используется в механизме работы собачьих ультразвуковых свистков). А вот домашние кошки слышат почти, как мы. Китообразные, с другой стороны, испускают и воспринимают звуки намного более высокочастотные – до 100 кГц. Такие звуки могут использоваться в социальной жизни вида, а у некоторых видов, например, дельфина, и в эхолокации. Интенсивность и продолжительность эмиссии звука у них возрастает по мере приближения к препятствию или при поиске пищи.
|
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 2222; Нарушение авторских прав?; Мы поможем в написании вашей работы!