КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Часть 5. Обоняние у позвоночных
|
|
|
|
Носовая полость большинства дышащих воздухом позвоночных содержит по крайней мере два различных обонятельных органа: первый и наиболее важный – это основной обонятельный эпителий, покрывающий носовые проходы, и второй – вомероназальный орган, которые будут рассмотрены ниже.
- Обонятельный эпителий полости носа
У высших приматов, включая человека, обонятельный эпителий сравнительно невелик. Этому можно искать объяснение в том, что запахи под пологом леса разносятся недалеко. Но главное, приматы – животные с дневной активностью, и полагаются преимущественно на зрение при определении расстояний и размеров. У ночных форм животных, которым от зрения намного меньше пользы, развивались и достигли совершенства и доминируют два других дистантных чувства – слух и обоняние. Размеры обонятельного эпителия у млекопитающих варьируют от 2-4 см2 (человек) и 9,3 см2 (кролик) до 18 см2 (собака) и 21 см2 (домашняя кошка). Надо отметить, однако, что эти цифры не дают точного представления об «остроте» обоняния, поскольку здесь не учтено число обонятельных рецепторных клеток на единицу поверхности.
У млекопитающих обонятельный эпителий развился на тонких пластинчатых костях полости носа – носовых раковинах. Эта область выполняет двойную функцию – не только подставляя обонятельный эпителий под поток поступающего воздуха и создавая сопротивление его потоку, но и нагревая воздух перед его поступлением в легкие. Наличие или отсутствие носовых раковин в отложениях костных остатков используется для определения того, был ли их давным-давно умерший обладатель теплокровным. Обонятельный эпителий содержит три типа клеток:
Ø нейросенсорные – обонятельные рецепторные клетки,
Ø опорные клетки, секретирующие слизь – близкие по свойствам к глиальным клеткам,
Ø базальные, которые (подобно стволовым клеткам) способны делиться и формировать новые функциональные нейроны в течение жизни животного (рис. 51).
 |
Обонятельные клетки – это единственные нейроны у млекопитающих, способные обновляться в течение всей жизни животного.
Как уже было показано выше в слуховой, соматосенсорной и, в меньшей степени, вкусовой системах сенсорный эпителий регистрирует пространственную информацию. «Картирующая» функция в обонятельном эпителии не так выражена. В первом приближении клетки, специализированные к различению специфических запахов, распределены в случайном порядке, однако более пристальное изучение заставляет предположить, что и здесь 
 присутствуют зачатки пространственной организации. Картирование с помощью 2-деоксиглюкозы (которая дает возможность определить активные клетки) показало наличие групп клеток, связанных с определенным запахом. Например, бутанол возбуждает клетки передней области, тогда как лимонен активирует клетки задней части слизистой. Недавние исследования также заставляют предположить, что рецепторные клетки организованы в передне-задние полосы, называемые зонами экспрессии, каждая из которых содержит полный набор клеток. Вероятно, существует три не перекрывающиеся зоны экспрессии, которые перекрываются с меньшей, четвертой зоной.
присутствуют зачатки пространственной организации. Картирование с помощью 2-деоксиглюкозы (которая дает возможность определить активные клетки) показало наличие групп клеток, связанных с определенным запахом. Например, бутанол возбуждает клетки передней области, тогда как лимонен активирует клетки задней части слизистой. Недавние исследования также заставляют предположить, что рецепторные клетки организованы в передне-задние полосы, называемые зонами экспрессии, каждая из которых содержит полный набор клеток. Вероятно, существует три не перекрывающиеся зоны экспрессии, которые перекрываются с меньшей, четвертой зоной.
- Обонятельные рецепторные клетки
Как показано на рис. 51, обонятельные рецепторные клетки (ОРК) – биполярные с одним неветвящимся дендритом, который проходит между опорными клетками, заканчивающийся небольшим вздутием – обонятельной булавой. Из нее вырастают до 20 длинных ресничек. Эти реснички представляют собой сенсорную поверхность обонятельной клетки. Обычно они погружены в слой слизи, покрывающей эпителий, образуя с ним плотный матрикс. Обонятельная клетка имеет двойную функцию – восприятие стимула и передачу нервного импульса в мозг, а потому является нейросенсорной клеткой, ее иногда называют сенсорным нейроном.
Слизь, в которой лежат обонятельные реснички, содержит большое количество небольших (20 кДа) белков, во многих отношениях похожих на одорант-связывающие белки (ОСБ) сенсилл насекомых. Они секретируются железами слизистой носа и встречаются не только в слизи, покрывающей обонятельный эпителий, но и в слизи, покрывающей чисто респираторный эпителий. Этот факт, а также их сравнительно небольшой размер, позволяют предположить, что эти белки не играют столь же важной роли, как ОСБ насекомых и бактерий. Если эта роль и существует, то сродство этих белков к молекулам-одорантам очень неселективно, а функция сводится к связыванию одорантов и обеспечению их взаимодействия с рецепторными клетками.
Ультраструктура обонятельных ресничек не сильно отличается от таковой других ресничек. Они содержат обычную аксонему, которая, по крайней мере у млекопитающих, неподвижна. Эти реснички необычно длинные и тонкие – от 5 до 250 мкм в длину и лишь 100-250 нм в диаметре.
Пучок (от 5 до 40) длинных тонких ресничек, выходящий из булавы обонятельной клетки, увеличивает сенсорную поверхность. Электронная микроскопия замороженных сколов показывает, что мембрана каждой реснички содержит большое количество глобулярных частиц – предполагается, что это обонятельные рецепторные молекулы.
Обонятельная система способна детектировать и различать огромное количество запахов. Аналогично, как было показано на вкусовых клетках, остается предметом противоречий тот факт, является ли отдельная ресничка «генерализатором» или «специалистом», иными словами, отвечает ли ресничка на многие типы молекул пахучих веществ или только на один. В последние годы накапливаются данные в пользу их специализации. Но остается фактом, что независимо от генерализации или специализации, чувствительность их очень велика – даже человек способен обнаруживать пахучие вещества при концентрации 10-4М - 10-13М. Одним из признаков простуды является значительное снижение такой тонкой чувствительности. Это происходит потому, что обонятельные реснички оказываются погруженными в слишком большом слое слизи, вырабатываемой опорными клетками.
- Особенности биохимические реакций при передаче сигнала по обонятельным рецепторным клеткам
Обонятельные рецепторные белки. В последние годы идентифицировано большое семейство генов (несколько сотен, возможно, более тысячи), кодирующих обонятельные рецепторные белки, что составляет 1-2% генома. Это семейство генов кодирует семидоменные рецепторы.
Семидоменные обонятельные рецепторные молекулы отличаются от прочих членов надсемейства семидоменных рецепторов большим аминокислотным разнообразием в трансмембранных доменах 3,4 и, особенно, 5. Эти домены образуют существенную часть погруженной в мембрану «бочки» семидоменного рецептора. Поскольку похоже, что 
 именно внутренняя часть «бочки», образуемой этими доменами, формирует сайт, распознающий пахучие молекулы.
именно внутренняя часть «бочки», образуемой этими доменами, формирует сайт, распознающий пахучие молекулы.
Рассмотрим, как разные рецепторные молекулы приспособлены к распознаванию разных одорантов. Следует помнить, что в отличие от большинства других семидоменных рецепторных белков, сопряженных с G-белками, обладающих довольно избирательным сродством, – обонятельные рецепторные белки считаются в той или иной степени аффинными к целому ряду стереохимически сходных пахучих молекул. Есть данные, что обонятельные реснички специализированы, т.е. они способны экспрессировать только один тип обонятельных рецепторных молекул. Это однако не означает, что они реагируют только на один тип одорантов. Показано, что каждый данный рецептор стимулируется небольшим числом химически сходных молекул одорантов. С другой стороны, каждое пахучее вещество взаимодействует с некоторым числом разных рецепторов. Вероятно каждый обонятельный стимул взаимодействует со специфической группой рецепторных молекул, и этот набор рецепторов определяет индивидуальность запаха.
В то же время ситуация в слизистом слое сложна и динамична; небольшим изменениям химической структуры молекулы одоранта соответствует изменение набора стимулируемых рецепторов. Изменения концентрации молекул одоранта могут влиять также на число стимулируемых рецепторов и, таким образом, изменять информацию, передаваемую в обонятельную луковицу. Параллельно меняется и субъективное восприятие запаха.
Например, замещение гидроксильной группы октанола (органическое вещество, относится к классу жирных спиртов, содержится в эфирных маслах цитрусовых) на карбоксильную, казалось бы незначительное изменение, приводит к значительному изменению обонятельного восприятия запаха – вместо запаха, напоминающего апельсин и розу, появляется запах прогорклости и пота.
Некоторые пахучие вещества меняют качество своего запаха при изменении концентрации. Индол, например, имеет приятный цветочный запах при большом разведении и отвратительный гнилостный – в высокой концентрации.
Мембранная биохимия. Семидоменные рецепторы обонятельных ресничек связаны G-белковой системой с мембранно-связанными эффекторами. В большинстве случаев эффектором является аденилатциклаза (АЦ). Кроме того, это могут быть ФЛС-β, ИТФ и ДАГ.
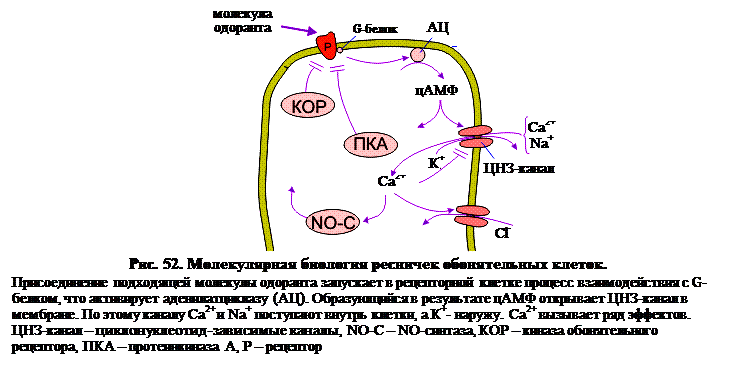
В тех случаях, когда вторичным мессенджером является цАМФ, он воздействует на каналы с малым диаметром поры (рис. 52). Поскольку каналы контролируются циклическими нуклеотидами, они и именуются циклонуклеотид-зависимыми (ЦНЗ-каналы). Хотя обонятельные ЦНЗ-каналы были выделены не первыми (эта честь принадлежит каналам палочек), к настоящему времени они охарактеризованы лучше других.
ЦНЗ-каналы. Выделение циклонуклеотид-зависимых каналов (ЦНЗ-каналов) было произведено следующим образом:. белок 63 кДа был выделен из наружных сегментов палочек и колбочек быка, при встраивании этого белка в искусственный липидный бислой можно регистрировать ток через единичный канал в ответ на цГМФ.
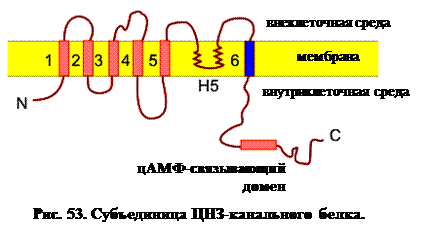
Чтобы определить, какие участки аминокислотной цепи локализованы в мембране, использовалась методика т.н. гидропатического анализа. В сущности, это означает просто проверку того, какие участки цепи состоят преимущественно из гидрофобных аминокислот. При анализе ЦНЗ-каналов выявлено шесть гидрофобных сегментов, т.е. канал имеет шесть трансмембранных сегментов (рис. 53). Такая молекулярная структура напоминает многие потенциал-заивисимые каналы: К+, Na+, Са2+, Сl- и т. д. Более детальный анализ показал, что четвертый трансмембранный сегмент S4 строго гомологичен сенсору потенциала S4 потенциал-зависимых каналов. Здесь присутствует также т.н. β-шпилька (петля ДНК) между сегментами S5 и S6. Следует отметить, что похожий домен Н5 присутствует в потенциал-зависимых каналах, где, как предполагается, он контролирует тип иона, способного пройти через канал. В отличие от потенциал-зависимых каналов, ЦНЗ-каналы имеют большой С-концевой цитоплазматический домен, который связывается с вторичными мессенджерами – циклическими нуклеотидами. Предполагается, главным образом по аналогии с хорошо изученным К+-каналом, что четыре ЦНЗ-канальных белка, показанных на рис. 53, образуют полный канал. Образован ли канал тетрамером одинаковых ЦНЗ-субъединиц (гомо-олигомер) или (как К+-канал) различными субъединицами ЦНЗ-тетрамера (гетеро-олигомер), пока неизвестно.
Сенсорная адаптация и градуальный ответ. Одной из интересных черт изолированных ЦНЗ-каналов обонятельных ресничек является то, что они не десенситизируются. Это означает, что при повторных предъявлениях одоранта ЦНЗ-каналы не утрачивают чувствительности. Сенсорная адаптация в обонятельных рецепторных клетках, тем не менее, имеет место. Вероятно, это происходит вследствие потока внутрь клетки ионов Са2+, которые закрывают ЦНЗ-каналы либо непосредственно (рис. 52), либо через активацию кальмодулина. Поток Са2+ внутрь клетки также ведет к десенситизации семидоменной рецепторной молекулы. Градуальность ответа на изменение концентрации одоранта связана с тем, что система вторичных мессенджеров цАМФ увеличивает (или уменьшает) число ЦНЗ-каналов в открытом состоянии.
 |
Чтобы различить запахи в реальном (биологическом) времени, ответ должен быть достаточно быстрым. В эксперименте показано, что пик продукции цАМФ наступает через 40-75 мс после аппликации одоранта и падает до нуля через 100-500 мс. Усиление сигнала G-белковым каскадом обеспечивает активацию множества каналов одним импульсом одоранта. Было показано, однако, что кинетика каналов достаточно медленна – открытое состояние может отставать от импульса цАМФ на несколько сот миллисекунд. Продолжительная активация семидоменных рецепторов постоянными концентрациями одоранта, таким образом, обеспечивает поддержание ЦНЗ-каналов в постоянно открытом состоянии импульсами цАМФ.
Аносмии. Генетический анализ обонятельной системы человека выделил несколько дюжин специфических аносмий (отсутствий обоняния). Например, один человек из десяти неспособен определить запах цианида, а один из тысячи – нечувствителен к бутилмер-каптану – активному агенту зловония, испускаемого скунсом. Эти аносмии возникают, вероятно, вследствие дефектов специфических обонятельных рецепторных молекул. Многие из них наследуются в соответствии с законами Менделя. Очевидна аналогия с цветовой слепотой у человека, которая также возникает в результате дефекта рецепторного белка, в последнеми случае – йодопсинов. В отличие от генетики аносмий, генетика цветовой слепоты сравнительно хорошо известна.
- Центральная обработка обонятельной информации
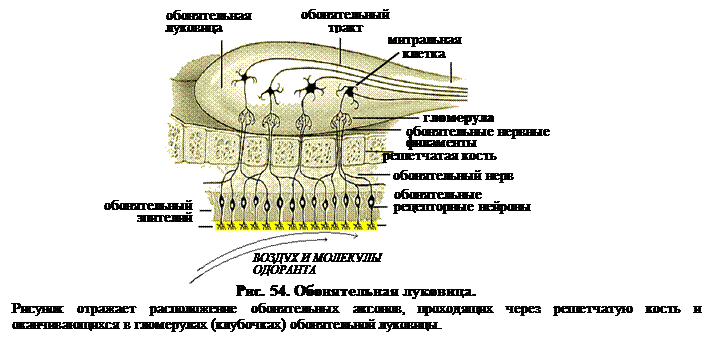
Обонятельные клетки являются нейросенсорными. Когда генераторный потенциал достигает порога, инициируется потенциал действия в начальном сегменте аксона. Рис. 54 показывает, что эти аксоны проходят через отверстия решетчатой кости и входят в обонятельную луковицу, лежащую прямо над ней.
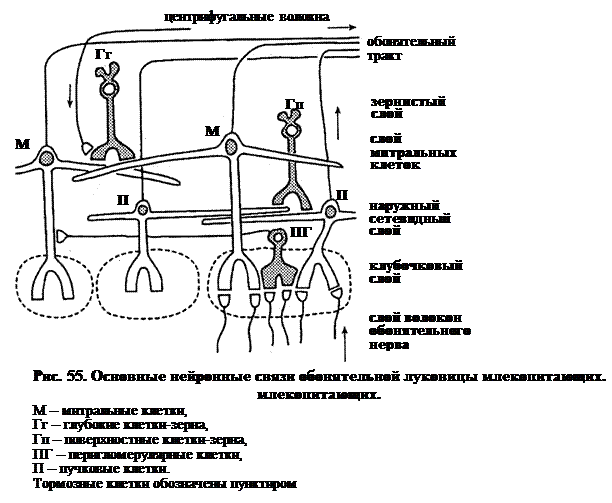
Аксоны обонятельных рецепторных клеток оканчиваются сложными синаптическими контактами с дендритами митральных (названных так, потому что первым микроскопистам их тела напомнили епископскую митру), пучковых и перигломерулярных клеток.
В обонятельной луковице кролика конвергенция составляет 1000 к 1, т.е. около 1000 обонятельных аксонов конвергируют на разветвлениях апикально дендрита одной митральной клетки. Около 25 таких дендритов вместе с терминалями аксонов формируют сферическое образование, именуемое гломерулой (обонятельный клубочек). Т.о., на одной гломеруле конвергирует около 25 000 обонятельных аксонов. В обонятельной луковице кролика около 2000 гломерул. Между гломерулами расположены короткоаксонные перигломерулярные клетки. Аксоны этих клеток соединяют их с одной гломерулой, а дендритное дерево – с соседней. Есть надежные данные, говорящие о том, что перигломерулярные клетки являются тормозными.
 |
Глубже в обонятельной луковице (рис. 55) образуют слой тела митральных клеток. Между ними лежат тела клеток-зерен. Клетки-зерна лежат глубже тонкого митрального слоя и заполняют центральную часть луковицы. Зерна, как и перигломерулярные клетки, являются тормозными. Такая стратификация клеточной организации обонятельной луковицы демонстрирует сходство с сетчаткой. Основа вертикальной передачи информации – это аксоны митральных и, в меньшей степени, пучковых клеток, которые проходят два уровня горизонтальной модуляции.
Детальная электронная микроскопия обонятельной луковицы показывает сложность синаптических контактов. Нейрохимия их не менее сложна. Здесь идентифицировано около 12 медиаторов, включая ацетилхолин, дофамин, γ-аминомасляную кислоту (ГАМК) и несколько нейроактивных пептидов.
 |
В последнее время собрано все больше подтверждений тому, что обонятельные рецепторные клети, специализированные реагировать на одни и те же запахи, проецируются в одни и те же гломерулы. Т.е., ОРК, отвечающие на небольшой класс сходных пахучих веществ или единственное вещество, могут быть распределены сравнительно беспорядочно в зонах экспрессии обонятельного эпителия, их аксоны конвергируют на небольшом числе гломерул или даже на одной (рис. 56). Более того, анатомическое расположение таких гломерул идентично у разных животных одного и того же вида. Таким образом, данный запах вызывает сходные паттерны активности в гломерулярной популяции обонятельной луковицы у всех представителей данного вида. Поскольку разные компоненты запаха активируют различные популяции ОРК, их совокупность является сложной и уникальной. Удивительно то, что запахи, как и афферентация других дистантных чувств, в центральной нервной системе могут кодироваться пространственной картиной распределения возбуждения в соответствующей проективной зоне.
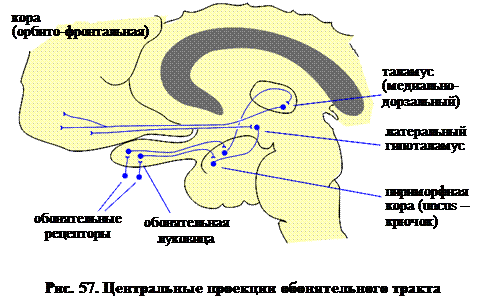
Из луковицы обонятельная информация передается в мозг аксонами митральных (и пучковых) клеток, образующих обонятельный тракт. Как показывает рис. 57, обонятельный тракт проходит под лобной долей и оканчивается в одной из эволюционно наиболее древних частей коры – пириформной доле (или uncus). Следующий нейрон передает обонятельную информацию в таламус, а последнее волокно передает ее в орбито-фронтальную новую кору. Помимо этого таламического пути, группа волокон проецируется через uncus в латеральный гипоталамус и отсюда в орбито-фронтальную новую кору. Возможно, именно этот путь запускает эмоциональные реакции, которые вызывают знакомые запахи и ароматы.
 |
- Вомероназальный орган позвоночных и феромоны
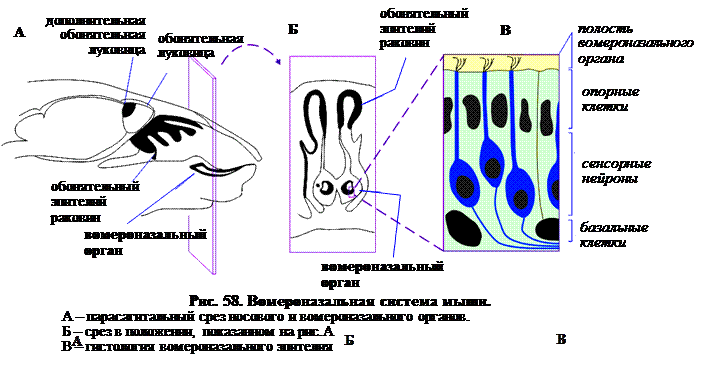
Положение вомероназального органа у мыши показано на рис. 58. Это парный слепой канал под носовой полостью. Он развивается у воздуходышащих наземных четвероногих, включая человека, и отсутствует у рыб. В нем присутствует обонятельный эпителий, который на клеточном уровне похож на тот, что покрывает раковины полости носа. Обонятельные рецепторные клетки (ОРК) – нейросенсорные клетки – поддерживаются эпителиальными и базальными клетками эпителия. ОРК, как и в носовой полости, регенерируют в течение всей жизни, но вместо ресничек у ОРК развиваются микроворсинки («микровиллы»).
 |
Молекулярная биология обонятельных рецепторных клеток вомероназального органа существенно отличается от ОРК носовых ходов. Как и другие ОРК, они имеют полный набор семидоменных рецепторных молекул, но их аминокислотная последовательность не имеет сходства с рецепторами других ОРК. Повидимому, эти клетки имеют свою долгую независимую историю. Недавно было показано, что в слизистой вомероназального органа присутствуют два различающихся семейства семидоменных рецепторов, каждое из которых включает от 100 до 200 генов и развивалось, по-видимому независимо. Для представителей одного из них типичен длинный внеклеточный N-концевой домен, сходный с метаботропным глютаматным рецептором (mGluR). G-белковая сигнальная система, представленная в ОРК носовых ходов, в клетках вомероназального органа не экспрессируется. В отличие цАМФ-зависимой системы носовой полости, в вомероназальных ОРК действует система ИТФ. Это отличие, в дополнение к несходству семидоменных рецепторных белков, наталкивает на мысль, что вомероназальная и носовая обонятельные системы возникли в эволюции независимо. Поскольку у рыб вомероназальный орган не обнаружен, возможно, что исходно два этих типа эпителия были перемешаны, а разделились только в ходе эволюции назменых позвоночных.
Змеи пользуются вомероназальным органом при преследовании жертвы. То, что вомероназальный орган участвует в детектировании феромонов позвоночных, подозревали уже давно. Самый яркий пример восприятия феромонов у млекопитающих получен в исследованиях полового поведения золотистых хомячков. Садка самца запускается феромонами из вагинального секрета самок. Включение полового созревания ускоряется мочой самцов и тормозится мочой самок. Предполагается, что эти и другие запускаемые феромонами поведенческие и физиологические реакции опосредованы вомероназальным органом. Исследования в настоящее время развиваются, и специфические молекулы феромонов еще предстоит охарактеризовать.
Афферентные волокна из вомероназального эпителия проецируются в дополнительную обонятельную луковицу, у большинства млекопитающих расположенную кзади от каудальной по отношению к основной обонятельной луковице (рис. 58). Интересно отметить, что, как и обонятельный эпителий носовых ходов, вомероназальный эпителий подразделяется на зоны. Это подразделение обозначено экспрессией разных G-белков в апикальной и базальной частях органа. Эти зоны сохраняются и в проекциях в дополнительную обонятельную луковицу. Апикальные ОРК проецируются в переднюю, а базальные – в заднюю часть луковицы. Нервные окончания находятся в гломерулах, которые, однако, менее выражены, чем в основной обонятельной луковице. Вероятно, между луковицами есть существенная разница в представительстве. В отличие от пространственной карты, характерной для основной луковицы, в дополнительной луковице представительство более сложное и мозаичное. Его изучение – это предмет исследований, использующих все современные методики молекулярной нейробиологии. Предполагается, что сложная природа представительства может быть связана с биологической функцией системы. Возможно, дополнительная луковица призвана реагировать исключительно на особые видоспецифические сочетания веществ соответствующего феромона и просто игнорировать все остальные. Проекции из дополнительной луковицы отличаются от таковых основой луковицы. Вместо проекций в кору, дополнительная луковица связана исключительно с лимбической системой – миндалиной и гипоталамическими ядрами. Известно, что эти ядра играют важную роль в половом и репродуктивном поведении.
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 1973; Нарушение авторских прав?; Мы поможем в написании вашей работы!