КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Инфекционный процесс. Факторы, обуславливаемые его
|
|
|
|
Класс VII: вирусы, содержащие двуцепочечную ДНК, реплицирующиеся через стадию одноцепочечной РНК
Класс V: вирусы, содержащие одноцепочечную (−)РНК
Класс IV: вирусы, содержащие одноцепочечную (+)РНК
Класс III: вирусы, содержащие двуцепочечную РНК
Класс II: вирусы, содержащие одноцепочечную ДНК
Класс I: вирусы, содержащие двуцепочечную ДНК
Методы выделения чистой культуры облигатных анаэробов
I этап — обогащение на среде Китт — Тароцци, предварительно прокипяченной в течение 30 минут. После посева среду прогревают 15 минут при 80°С для уничтожения вегетативных форм, споры анаэробов при этом сохраняются. II этап — получение изолированных колоний На средах Китт — Тароцци обнаруживается помутнение с пузырьками газа. Выделение чистой культуры проводится по одному из следующих методов:
А) по Цейсслеру — каплю материала со среды Китт — Тароцци засевают в чашку с кровяным агаром и распределяют материал шпателем, этим же шпателем производят посев во второй и третьей чашках;
Б) по Вейнбергу: каплю материала со среды Китт — Тароцци переносят в растопленный и слегка остуженный сахарный агар, затем набирают в трубки Виньял — Вейона и инкубируют в термостате. Ш этап — выделение чистой культуры на среде Китт — Тароцци.
Вирусы, содержащие двуцепочечную ДНК для репликации попадают в ядро клетки, так как им требуется клеточная ДНК-полимераза. Также репликация ДНК этих вирусов сильно зависит от стадии клеточного цикла. В некоторых случаях вирус может вызывать деления клетки, что может приводить к раковому перерождению. Примерами таких вирусов являются Herpesviridae, Adenoviridae и Papovaviridae.
У представителей семейства Poxvirus геномная ДНК реплицируется не в ядре.
Вирусы семейств Circoviridae и Parvoviridae реплицируют геномную ДНК в ядре и в ходе репликации образуют интермедиат — двуцепочечную ДНК.
Как и большинство РНК-вирусов, представители класса III реплицируют геномную РНК в цитоплазме и используют полимеразы хозяина в меньшей степени, чем ДНК-вирусы. Класс III включает в себя два крупных семейства Reoviridae и Birnaviridae. Репликация моноцистронная, геном сегментирован, каждый ген кодирует один белок.
Непосредственно на (+) геномной РНК вирусов IV класса может идти синтез белка на рибосомах клетки хозяина. Вирусы классифицируют на две группы, в зависимости от особенностей РНК:
у вирусов с полицистронной мРНК трансляция приводит к образованию полипротеина, который затем разрезается на зрелые белки. С одной цепи РНК может синтезироваться несколько разных белков, что снижает длину генов.
вирусы со сложной транскрипцией содержат субгеномные мРНК, синтез белка идет со сдвигом рамки считывания, также используется протеолитический процессинг полипротеинов. Эти механизмы обеспечивают синтез разных белков с одной цепи РНК.
Геномные РНК вирусов класса V не могут быть транслированы на рибосомах клетки хозяина, предварительно требуется транскрипция вирусными РНК-полимеразами в (+)РНК. Вирусы пятого класса классификации по Балтимору классифицируют на две группы:
вирусы, содержащие несегментированный геном, на первом этапе репликации происходит транскрипция (−)РНК вирусной РНК-зависимой РНК-полимеразной в моноцистронную мРНК, и далее синтезируются дополнительные копии (+)РНК, служащие матрицами для синтеза геномных (−)РНК. Репликация геномных РНК таких вирусов осуществляется в цитоплазме.
вирусы с сегментированными геномами, репликация геномных РНК которых происходит в клеточном ядре, вирусная РНК-зависимая РНК-полимераза синтезирует моноцистронные мРНК с каждого сегмента генома. Наибольшим отличием данной группы вирусов от другой группы пятого класса состоит в том, что репликация осуществляется в двух местах.
Представители данного класса входят в состав семейств: Arenaviridae, Orthomyxoviridae, Paramyxoviridae, Bunyaviridae, Filoviridae и Rhabdoviridae.
Класс VI: вирусы, содержащие одноцепочечную (+)РНК, реплицирующиеся через стадию ДНК
Наиболее хорошо изученным семейством данного класса вирусов, являются ретровирусы. Вирусы класса VI используют фермент обратную транскриптазу для превращения (+)РНК в ДНК. Вместо использования РНК в качестве матрицы для синтеза белков, вирусы этого класса используют матрицу ДНК, которая встраивается в геном хозяина ферментом интегразой. Дальнейшая репликация происходит при помощи полимераз клетки хозяина. Наиболее хорошо изученным представителем данной группы вирусов является ВИЧ.
Небольшая группа вирусов, в состав которой входит вирус гепатита В, представитель семейства Hepadnaviridae, имеют двуцепочечную геномную ДНК, которая ковалентно замкнута в форме кольца и является матрицей для синтеза мРНК вируса, а также субгеномных РНК. Субгеномная РНК служит матрицей для синтеза ДНК-генома ферментом обратной транскриптазой вируса.
3. Морфология вирусов.
Вирусы относятся к царству Vira. Это мельчайшие микроорганизмы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только один тип нуклеиновой кислоты (ДНК или РНК). Они отличаются особым разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки и затем происходит их сборка в вирусные частицы. Вирусы, являясь облигатными внутриклеточными паразитами, размножаются в цитоплазме или ядре клетки. Сформированная вирусная частица называется вирионом.
Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий.
Форма вирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги).
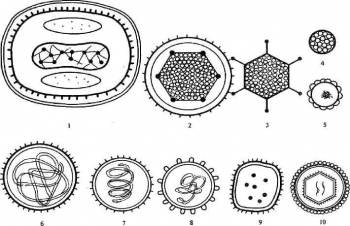
1 вирус оспы; 2 вирус герпеса; 3 аденовирус; 4 паповавирус; 5 гепаднавирус; 6 парамиксовирус; 7 вирус гриппа; 8 коронавирус; 9 аренавирус; 10 ретровирус;
Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одними из самых мелких вирусов являются вирусы полиомиелита и ящура (около 20 нм), цирковирусы (16 нм), наиболее крупным вирус натуральной оспы (около 350 нм). Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, то есть имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными.
Различают просто устроенные (например, вирус полиомиелита) и сложно устроенные (например, вирусы гриппа, кори) вирусы. У просто устроенных вирусов нуклеиновая кислота связана с белковой оболочкой, называемой капсидом (от лат. capsa футляр). Капсид состоит из повторяющихся морфологических субъединиц капсомеров. Нуклеиновая кислота и капсид, взаимодействуя друг с другом, образуют нуклеокапсид. У сложно устроенных вирусов капсид окружен дополнительной липопротеидной оболочкой суперкапсидом (производное мембранных структур клетки-хозяина), имеющим «шипы». Капсид и суперкапсид защищают вирионы от влияния окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с клетками, определяют антигенные и иммуногенные свойства вирионов. Внутренние структуры вирусов называют сердцевиной.
Для вирионов характерен спиральный, кубический и сложный типы симметрии капсида. Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида, кубический образованием изометрического полого тела из капсида, содержащего вирусную нуклеиновую кислоту.
Кроме обычных вирусов, известны и так называемые неканонические вирусы прионы белковые инфекционные частицы, имеющие вид фибрилл размером 10-20 х 100 200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирусной инфекции (болезни Крейтцфельдта Якоба, куру и др.). Другими необычными агентами, близкими к вирусам, являются вироиды небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белка, вызывающие заболевания у растений.
Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса).
По форме вирионы могут быть: округлыми, палочковидными, в виде правильных многоугольников, нитевидными и др.
Размеры их колеблются от 15–18 до 300–400 нм.
В центре вириона – вирусная нуклеиновая кислота, покрытая белковой оболочкой – капсидом, который имеет строго упорядоченную структуру. Капсидная оболочка построена из капсомеров.
Нуклеиновая кислота и капсидная оболочка составляют нуклеокапсид.
Нуклеокапсид сложноорганизованных вирионов покрыт внешней оболочкой – суперкапсидом.
Вирусные белки подразделяют на:
1) геномные – нуклеопротеиды. Обеспечивают репликацию вирусных нуклеиновых кислот и процессы репродукции вируса;
2) белки капсидной оболочки – простые белки, обладающие способностью к самосборке. Они складываются в геометрические структуры, в которых различают несколько типов симметрии: спиральный, кубический или смешанный;
3) белки суперкапсидной оболочки – это сложные белки. Выполняют защитную и рецепторную функции.
Среди белков суперкапсидной оболочки выделяют:
а) якорные белки (обеспечивают контакт вириона с клеткой);
б) ферменты (могут разрушать мембраны);
в) гемагглютинины (вызывают гемагглютинацию);
г) элементы клетки хозяина.
Существует четыре типа взаимодействия:
1) продуктивная вирусная инфекция (происходит репродукция вируса, а клетки погибают);
2) абортивная вирусная инфекция (репродукции вируса не происходит, а клетка восстанавливает нарушенную функцию);
3) латентная вирусная инфекция (идет репродукция вируса, а клетка сохраняет свою функциональную активность);
4) вирус-индуцированная трансформация (клетка, инфицированная вирусом, приобретает новые, свойства).
4. Структуры простых и сложных вирусов.
Различают простые и сложные вирионы. Простые вирионы состоят из нуклеиновой кислоты, окруженной снаружи белковой оболочкой, которую называют капсидом (ящиком, футляром), сложные — имеют дополнительную внешнюю оболочку (суперкапсид, пеплос).
Структура и химический состав простых вирионов. В состав простых вирусов, типичным представителем которых является вирус табачной мозаики, входят только капсидные белки, но у некоторых из них содержатся также геномные, или терминальные, ковалентно связанные с концом вирионной нуклеиновой кислоты и участвующие в регуляции вирусного генома.
Капсидные белки простоорганизованных вирионов обычно состоят из 1-3 вирусоспецифических белков (полипептидных цепей). При этом каждый из них обладает способностью к самосборке, в начале которой из идентичных полипептидных цепей образуются отдельные структурные элементы (субъединицы) или капсомеры белкового чехла, вслед за чем при их взаимодействии с нуклеиновой кислотой вириона происходит полное самопроизвольное формирование капсида. Количество капсомеров в капсиде вирусов варьирует от трех-шести десятков до многих сотен. У одних вирусов капсомеры имеют овальную или округлую форму, у других - пяти- или шестигранную.
По характеру расположения капсомеров вирусы делят на три группы: с кубическим, спиральным и смешанным типом симметрии. Большинство патогенных для человека вирусов имеет кубический тип симметрии. Спиральный тип симметрии нередко встречается среди вирусов растений. Смешанный тип симметрии для простых вирусов нетипичен.
Структура и химический состав сложных вирионов. Сложно устроенные вирусы в капсиде имеют много разновидностей белков. Кроме капсидных и геномных белков могут также содержать ферменты, участвующие в репликации и транскрипции вирусного генома, например ДНК- и РНК-полимеразы. Формирование их капсидов и нуклеокапсидов происходит тоже в процессе самопроизвольной сборки, но цикл полного образования сложных вирионов носит многоступенчатый характер.
В суперкапсиде сложных вирусов, представляющем собой липидный бислой, в котором расположены пепломеры, превалируют гликопротеиды. Являясь типичными внутримембранными белками, они у большинства вирусов образуют поверхностные «шипы», длина которых достигает 5-10 нм. Чаще всего шипы построены из нескольких молекул идентичного белка, и те вирусы, которые имеют один гликопротеид, естественно, обладают одним типом шипов, при наличии в них двух гликопротеидов - двумя типами. Есть, однако, вирусы, имеющие 2-3 гликопротеида, формирующие один тип шипов.
Суперкапсидные вирусные гликопротеиды выполняют две основные функции: 1) распознают специфические клеточные рецепторы и взаимодействуют с ними, что дало повод называть их прикрепительными белками, и 2) обусловливают проникновение вируса в клетки, инициируя слияние его оболочек с клеточными мембранами, вследствие чего их называют белками слияния.
Количество углеводов в гликопротеидах может достигать 10% общей массы вириона. Обычными сахарными остатками в них являются сахароза, фруктоза, манноза, галактоза, нейраминовая кислота. Углеводы гликопротеидов обеспечивают сохранение конформации белка и его устойчивость к протеазам.
5. Строение вирусных геномов.
Все вирусные геномы являются гаплоидными, т.е. содержат одну копию каждого гена. Исключение составляют ретровирусы, которые обладают диплоидным геномом. Геномы ДНК-вирусов позвоночных представлены одной двуспиральной молекулой за исключением парво- и цирковирусов.
Все РНК-вирусы позвоночных за исключением рео- и бирнавирусов имеют одноцепочечные геномы. Геном некоторых РНК-вирусов состоит из нескольких (2-12) уникальных фрагментов, каждый из которых кодирует, как правило, один белок. РНК-вирусы с односпиральным геномом могут иметь различную полярность. Если они имеют ту же полярность, что и мРНК, то они могут прямо индуцировать синтез вирусного белка и считаются положительно (+) полярными.
Если геномная нуклеотидная последовательность комплементарна мРНК, то они считаются отрицательно (—) полярными. К ним относятся: парамиксо-, рабдо-, фило-, ортомиксо-, арена- и буньявирусы. Все они имеют вирионную РНК-зависимую полимеразу (транскриптазу), которая в инфицированной клетке транскрибирует положительно-полярную РНК на матрице геномной вирусной РНК. У аренавирусов, по крайней мере, у одного рода буньявирусов, один из РНК-сегментов является двуполярным. Обычно у (+)полярных РНК-вирусов З'-конец имеет polyA-последовательность, а 5'-конец имеет кэп-структуру.
Размер геномов РНК-вирусов (одноцепочечных 1,7—21 т.н.; двуцепочечных — 18—27 т.п.н.) значительно меньше размера генома многих ДНК-вирусов. Поэтому РНК-вирусы, как правило, кодируют меньше белков, чем ДНК-вирусы. Масса генома различных вирусов находится в пределах от 1 % (орто- и пара-миксовирусы) до 32% (парвовирусы) от массы вириона.
Число генов в вирусных геномах колеблется от 3-4 у самых простых вирусов до многих десятков у сложно устроенных. ДНК-содержащие вирусы так же, как про- и эукариоты, имеют структурные гены, кодирующие белки-ферменты, и регулятор-ные гены, детерминирующие образование репрессоров, подавляющих, в частности, функцию структурных.
Считывание информации с оперонов контролируется энхансером (англ. enhancer) или усилителем транскрипции; промотором (лат. promotum - продвигать), ответственным за ее инициацию (начало), с которым связывается фермент РНК-полимераза, осуществляющая транскрипцию ДНК; оператором (от лат. работник), регулирующим транскрипцию оперона (или отдельных генов) и терминатором (лат. terminare - ограничивать), прекращающим ее.
При этом регуляторные участки оперона представляют собой короткие последовательности нуклеотидов ДНК; энхансер, промотор и оператор расположены в его начале (перед структурными генами), а терминатор - в конце.
В структурных генах вирусных оперонов, как и в клетках эукариот, имеются кодируемые участки нуклеотидных последовательностей, несущих информацию (экзоны), и некодируемые вставочные последовательности (интроны), которые после транскрипции в процессе созревания (процессинга) иРНК вырезаются с одновременным считыванием экзонов, что называется сплайсингом (англ. splice - соединять, сращивать). Сплайсинг при формировании зрелых иРНК отмечается чаще всего среди вирусов, имеющих ядерную локализацию транскрипции, поскольку ферменты, осуществляющие его, находятся в ядре.
Вирусные иРНК в отличие от иРНК про- и эукариот могут направлять синтез не одного, а двух-трех белков, что позволяет им при минимальном содержании генетического материала увеличивать генетическую информацию. Достигается это двухкратным считыванием одной и той же иРНК с находящихся в ней в разных участках двух-трех иницирующих АУГ-кодонов. Происходит это, естественно, при трансляции белка и в каждом случае определяется, какой АУГ-кодон (первый или последующие) узнает малая рибосомная единица. Образующиеся полипептиды с разных инициирующих кодонов будут копиями, отличающимися разве только длиной.
При сдвиге рамки считывания на один или два нуклеотида и появлении нового генетического кода молекула иРНК может транслироваться с образованием таких полипептидов, у которых нет идентичных аминокислотных последовательностей, вследствие чего их называют уникальными белками.
Нередким способом увеличения генетической информации у вирусов является трансляция гигантских полипептидов-предшественников с последующим нарезанием их на более мелкие.
6. Капсиды вирионов.
Вирусная нуклеиновая кислота окружена однослойной или двойной белковой оболочкой (капсид). Нуклеиновая кислота вируса в сочетании с капсидом получила название нуклеокапсид. Вирусные капсиды состоят из небольших повторяющихся субъединиц (капсомеры), организованных в симметричные конструкции. Повторение субъединиц облегчает сборку вирусных белков до стадии зрелых вирионов и уменьшает количество генетической информации, необходимой для кодирования структурных протеинов. Капсиды формируются путем самосборки их структурных субъединиц.
Капсид выполняет несколько функций.
- Защита генетического материала (ДНК или РНК) вируса от механических и химических повреждений.
- Определение потенциала к заражению клетки.
- На начальных стадиях заражения клетки: прикрепление к клеточной мембране, разрыв мембраны и внедрение в клетку генетического материала вируса.
Существуют два основных варианта структурной симметрии капсида — икосаэдр и спираль. Некоторые из наиболее крупных вирусов, такие как поксвирусы, имеют более сложную структурную организацию. Капсид ретровирусов имеет икосаэдрическую симметрию, а их ядро — спиральную симметрию. Вирусы, чьи капсиды имеют икосаэдрическую симметрию, в целом подчиняются принципам физической организации, в соответствии с которыми общее количество структурных единиц ограничено. Нуклеиновая кислота вирусов, имеющих форму икосаэдра, находится обычно в сжатом состоянии и геометрически независима от структуры окружающего капсида.
В случае спиральной симметрии (например, у вируса табачной мозаики) составные части капсида формируют цилиндр из уложенных по спирали белковых глобул, внутри которого находится генетический материал вируса. В случае икосаэдрической симметрии (например, у многих бактериофагов) образуется квази-сферическая структура капсида. В случае «закутанного» капсида отдельные части капсида (спирального или икосаэдрического) открыты окружающей
среде, в то время как большая часть его скрыта мембраной.
Вирионы вируса табачной мозаики имеют форму палочек диаметром 15—17 нм и длиной до 300 нм. Методом рентгеноструктурного анализа было показано, что в капсиде вируса табачной мозаики на каждые три витка спирали приходится 49 белковых субъединиц. Каждая субъединица представлена молекулой белка с молекулярной массой 17 400. Капсид вируса табачной мозаики насчитывает 130 витков. Внутри капсида образуется полый канал диаметром 4 нм. Генетическим материалом вируса табачной мозаики является одноцепочечная РНК, плотно уложенная в желобке спирального капсида. Нуклеотиды РНК контериофаги по форме делятся на четыре группы: фаги, имеющие кубическую головку и отросток; кубические фаги; нитевидные фаги; плеоморфные фаги.
7. Строение внешней оболочки суперкапсида вириона.
Одетые вирусы. Некоторые вирусы могут содержать поверх капсида особую оболочку — суперкапсид, организованный двойным слоем липидов и специфичными вирусными белками, наиболее часто образующими выросты-шипы, пронизывающие липидный бислой. Такие вирусы называют «одетыми».
Образование суперкапсида происходит на поздних этапах репродуктивного цикла, обычно при отпочковывании дочерних популяций.
Липиды. Основная функция липидов — стабилизация структуры вирусов. Деградация или утеря липидов приводит к потере инфекционных свойств, так как такие вирусные частицы теряют стабильность своего состава и, соответственно, способность к заражению клеток. Состав липидов обычно зависит от характера «почкования» вирусной частицы. Например, у вируса гриппа состав липидного бислоя аналогичен таковому в клеточных мембранах. Герпес-вирусы почкуются через ядерную мембрану, поэтому набор липидов суперкапсида отражает состав липидов ядерной мембраны. Гликопротеины входят в состав поверхностных структур суперкапсида (например, «шипов»). Сахара, входящие в состав гликопротеинов, обычно происходят из клетки-хозяина. Поверхностные белки «голых» вирусов обеспечивают взаимодействие вирусов с клеточными рецепторами и последующее проникновение в клетку путём эндоцитоза. Большинство «одетых» вирусов имеют поверхностные специальные F-белки [от лат. fusio, слияние], обеспечивающие слияние вирусных суперкапсидов и клеточных мембран. Поверхностные белки — важный компонент, облегчающий проникновение вирусов в чувствительные клетки. Их характерное свойство — способность связываться с рецепторами на поверхности эритроцитов и агглютинировать их. Способность к гемагглютинации широко используют для определения количества вирусов.
«Голые» вирусы. Вирусы, не имеющие суперкапсида, называют «голыми». Как правило, они резистентны к действию эфира и более устойчивы к денатурации.
8. Неструктурные вирусные белки.
Неструктурные белки изучены гораздо хуже, чем структурные, поскольку их выделяют не из очищенных препаратов вирусов, а из зараженных клеток, и возникают трудности в их идентификации и очистке от клеточных белков.
К неструктурным белкам относятся:
- предшественники вирусных белков, которые отличаются от других неструктурных белков нестабильностью в зараженной клетке в результате быстрого нарезания на структурные белки;
- ферменты синтеза РНК и ДНК (РНК- и ДНК-26 полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома;
- белки-регуляторы;
- ферменты, модифицирующие вирусные белки, например протеиназы и протеинкиназы.
Однако многие неструктурные белки при ряде вирусных инфекций еще не идентифицированы и функции их не определены.
9. Стадии взаимодействия вируса с клеткой: продуктивная, интегративная формы.
Известны следующие типы взаимодействий «вирус-клетка»: продуктивный (образуется дочерняя популяция ), интегративный (вирогения), абортивный (дочерняя популяция не образуется) и интерференция вирусов (инфицирование чувствительной клетки разными вирусами).
Продуктивное взаимодействие «вирус-клетка» чаще носит литический характер, то есть заканчивается гибелью и лизисом инфицированной клетки, что происходит после полной сборки дочерней популяции. Гибель клетки вызывают следующие факторы: раннее подавление синтеза клеточных белков, накопление токсических и повреждающих клетку вирусных компонентов, повреждение лизосом и высвобождение их ферментов в цитоплазму.
Интегративное взаимодействие, или вирогения, не приводит к гибели клетки. Нуклеиновая кислота вируса встраивается в геном клетки-хозяина и в последующем функционирует как его составная часть. Наиболее яркие примеры подобного взаимодействия — лизогения бактерий и вирусная трансформация клеток.
Абортивное взаимодействие не приводит к появлению дочерней популяции и происходит при взаимодействии вируса с покоящейся клеткой (стадия клеточного цикла G0) либо при инфицировании клетки вирусом с изменёнными (дефектными) свойствами. Следует различать дефектные вирусы и дефектные вирионы. Первые существуют как самостоятельные виды и функционально неполноценны, так как для их репликации необходим «вирус-помощник» (например, для репликации аденоассоциированного вируса необходимо присутствие аденовирусов). Вторые составляют дефектную группу, формирующуюся при образовании больших дочерних популяций (например, могут образовываться пустые капсиды либо безоболочечные нуклео-капсиды). Особая форма дефектных вирионов — псевдовирионы, включившие в капсид нуклеиновую кислоту клетки-хозяина.
Интерференция вирусов происходит при инфицировании клетки двумя вирусами. Различают гомологичную (при инфицировании клетки родственными вирусами) и гетерологичную (если интерферируют неродственные виды) интерференцию. Это явление возникает не при всякой комбинации возбудителей, иногда два разных вируса могут репродуцироваться одновременно (например, вирусы кори и полиомиелита). Интерференция реализуется либо за счёт индукции одним вирусом клеточных ингибиторов (например, ИФН), подавляющих репродукцию другого, либо за счёт повреждения рецепторного аппарата или метаболизма клетки первым вирусом, что исключает возможность репродукции второго.
10. Последствия интеграции сирусного генома с геномом клетки.
Интегративный тип взаимодействия (вирогения) характеризуется встраиванием нуклеиновой кислоты вируса в хромосому клетки. Встраивание характерна для вирусов: бактериофагов, опухолеродных (онкогенных) вирусов, некоторых инфекционных вирусов (вирус гепатита В, аденовирус, ВИЧ). Вирус попал в клетку путем слияния мембран, попадают 2-е нити РНК. 2-е нити не нужны. Вирусная РНКаза разрушает одну нить, осталась одна.
Вриус РНК зависимая ДНК полимераза на матрице РНК строит одну нить ДНК. ДНК-полимераза достраивает 2-ю нить, получаем нормальную 2-х нитчатую ДНК, которая отправляется в ядро. ДНК вируса встраивается –вирогения - идет латантная стадия, после чего репродуктивная стадия. Клеточная ДНК-зависимая РНК-полимераза начинает строить + нити РНК. В рибосомах образуются вирусные белки, сборка и почкование и вирус выходит захватив с собой компоненты клетки, в том числе компоненты клеточной мембраны, что подавляет иммунитет. Идет изменение генома вируса, т.е. мутации. Для встраивания с хромосомой клетки необходима кольцевая форма двунитчатой вирусной ДНК. ДНК вируса, находящаяся в составе хромосомы клетки, называется ДНК-провирусом. При делении клетки, сохраняющей свои нормальные функции, ДНК-провирус переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. ДНК-провирус несет дополнительную генетическую информацию, в результате чего клетки приобретают ряд но вых свойств. Так, встраивание может явиться причиной возникновения ряда аутоиммунных и хронических заболеваний, разнообразных опухолей. Под воздействием ряда физических и химических факторов ДНК-провирус может исключаться из клеточной хромосомы и переходить в автономное состояние, что ведет к репродукции вируса.
11. Особенности репликации ДНК и РНК содержащих в вирусах.
Отличие вирусов от других организмов заключается в двух особенностях: 1) вирусная частица (вирион) содержит только один вид нуклеиновых кислот — или ДНК, или РНК; 2) вирионы отличаются необычной для живых существ простотой организации — они не имеют собственного метаболизма, не содержат клеточных органелл, в том числе рибосом, и очень часто состоят только из нуклеиновой кислоты, заключенной в белковую оболочку. В связи с этим вирусы способны размножаться исключительно за счет использования метаболического аппарата другой клетки, т. е. они являются внутриклеточными паразитами.
Цикл размножения вируса начинается с его прикрепления к поверхности клетки. Вирион содержит специальные белки, узнающие определенные вещества мембраны клетки-хозяина; эти вещества называют рецепторами вируса. Например, бактериофаг Т4 прикрепляется только к клеткам Е. coli, полиовирус — к определенным клеткам человека, а также обезьян, вирус гриппа — к клеткам слизистой оболочки дыхательных путей. После прикрепления вирион проникает через мембрану внутрь клетки; иногда в клетку попадает только нуклеиновая кислота вириона. Затем с использованием аппарата клетки-хозяина начинается репликация вирусного генома и синтез вирусных белков; из них путем самосборки образуются новые вирионы, которые освобождаются из клетки, либо разрушая ее (лизис клеток), либо проходя через мембрану без разрушения клетки.
Многие вирусы в качестве генетического материала содержат ДНК, но есть группа вирусов, геном которых представлен рибонуклеиновой кислотой. Размеры генома вирусов невелики. Например, в ДНК бактериофага Т4 обнаружено 135 генов; из них 36 генов кодируют синтез разных белков, входящих в оболочку фага, а остальные — гены белков, обеспечивающих переключение аппарата клетки-хозяина на синтез компонентов вируса, а также гены белков, выполняющих вспомогательную роль при самосборке вирионов. Механизм репликации генома ДНК-содержащих вирусов принципиально не отличается от репликации ДНК других организмов. РНК-содержащие вирусы по механизму репликации генома делятся на две группы. В одну группу входят полиовирус, вирусы гриппа, бешенства, везикулярного стоматита, реовирусы, вирусы свинки, кори и др.
Первый этап репликации вирусов связан с проникновением вирусной нуклеиновой кислоты в клетку организма-хозяина. Этому процессу могут способствовать специальные ферменты, входящие в состав капсида или внешней оболочки вириона, причем оболочка остается снаружи клетки или вирион теряет ее сразу после проникновения внутрь клетки. Вирус находит подходящую для его размножения клетку, контактируя отдельными участками своего капсида (или внешней оболочки) со специфическими рецепторами на поверхности клетки по типу «ключ – замок». Если специфические («узнающие») рецепторы на поверхности клетки отсутствуют, то клетка не чувствительна к вирусной инфекции: вирус в нее не проникает.
Для того чтобы реализовать свою генетическую информацию, проникшая в клетку вирусная ДНК транскрибируется специальными ферментами в мРНК. Образовавшаяся мРНК перемещается к клеточным «фабрикам» синтеза белка – рибосомам, где она заменяет клеточные «послания» собственными «инструкциями» и транслируется (прочитывается), в результате чего синтезируются вирусные белки. Сама же вирусная ДНК многократно удваивается (дуплицируется) при участии другого набора ферментов, как вирусных, так и принадлежащих клетке.
Синтезированный белок, который используется для строительства капсида, и размноженная во многих копиях вирусная ДНК объединяются и формируют новые, «дочерние» вирионы. Сформированное вирусное потомство покидает использованную клетку и заражает новые: цикл репродукции вируса повторяется. Некоторые вирусы во время отпочковывания от поверхности клетки захватывают часть клеточной мембраны, в которую «заблаговременно» встроились вирусные белки, и таким образом приобретают оболочку. Что касается клетки-хозяина, то она в итоге оказывается поврежденной или даже полностью разрушенной.
11. Противовирусы, химикотерапия, лечение.
Противовирусная терапия, в отличие от антибактериальной, обладает значительно меньшим арсеналом лечебных препаратов. Эффективность многих противовирусных химических соединений и препаратов установлена в экспериментальных исследованиях и в результате многочисленных клинических испытаний. Однако лишь немногие из них разрешены для широкого практического применения.
Особенности течения вирусной инфекции предполагает следующие терапевтические положения:
препараты должны отличаться надежностью противовирусного действия при минимальном повреждающем воздействии на клетки макроорганизма;
методы применения противовирусных средств ограничены недостаточными знаниями их фармакокинетики;
эффективность противовирусных химиопрепаратов в конечном итоге во многом зависит от защитных сил организма, напряженности иммунитета;
для практической медицины фактически недоступны методы определения чувствительности вирусов к применяемым химиопрепаратам.
По химическому составу и механизму действия противовирусные средства разделяются на три группы: 1) химиопрепараты; 2) интерфероны; 3) индукторы интерферонов.
Химиопрепараты. К противовирусным химиопрепаратам относятся аномальные нуклеозиды, производные адамантана, синтетические аминокислоты, аналоги пирофосфата, тиосемикарбазоны и другие вируцидные препараты.
Классификация основных противовирусных химиопрепаратов приведена в ряде обобщающих эту проблему работ, которые использованы при составлении настоящего пособия.
Указанные в таблице средства в большинстве своем эффективны при лечении гриппа и герпетической инфекции. К ним, к сожалению, быстро формируется резистентность, что существенно влияет на результаты лечения. При некоторых вирусных инфекциях, в частности герпетической, с обнадеживающими результатами использован препарат растительного происхождения - флакозид. Это флавоноид, полученный из растений бархата амурского и бархата Лаваля.
Интерфероны. Интерфероны (ИФН) относятся к биологическим противовирусным неспецифическим средствам. Они представлены практически во всех клетках организма и направлены на подавление репликации вирусов, их элиминацию и санацию организма. Механизм противовирусного действия ИНФ связан с блокадой синтеза вирусоспецифических белков путем распознавания и дискриминации информационных РНК.
Препараты ИНФ делятся по составу на альфа-, бета- и гаммаинтерфероны, а по способу получения - на природные человеческие, лейкоцитарные (первого поколения) и рекомбинантные ИНФ (второго поколения).
Со времени появления ИНФ стали вызывать большой интерес как с точки зрения профилактики, так и лечения вирусных заболеваний.
Результаты исследований показали, что ИНФ эффективны для лечения вирусных гепатитов, герпеса, острых респираторных инфекций, ВИЧ-инфекций и некоторых других заболеваний (таблица 6). Есть основания полагать, что экзогенные интерфероны, кроме прямого противовирусного действия, оказывают положительное влияние на иммунную систему. Они, в частности, нормализуют регуляторные механизмы клеточного звена иммунитета, способствуют индукции клетками альфа и гамма интерферона.
Индукторы интерферона. Индукторы интерферона представляют собой весьма разнообразную группу, высоко- и низкомолекулярных природных и синтетических соединений, способных вызвать образование интерферона в организме больного. В настоящее время имеются данные о целесообразности использования в комплексной терапии следующих индукторов интерферона: флуореонов, аналогов госсипола, сополимеров пирана, неовир и др. при герпетическом заболевании глаз, гриппе, риновирусной инфекции и прочих заболеваниях.
Инфекционный процесс (инфП) — типовой патологический процесс, возникающий под действием микроорганизмов. Инфекционный процесс — комплекс взаимосвязанных изменений: функциональных, морфологических, иммунобиологических, биохимических и других, лежащих в основе развития конкретных инфекционных болезней (инфБ). ИнфБ по их распространённости устойчиво удерживают третье место в мире (после болезней ССС и онкологических заболеваний). Крупные эпидемии и пандемии инфБ уносили многие миллионы жизней: от эпидемии чумы в средние века погибла треть населения Европы; в XVII-XVIII веках натуральной оспой ежегодно заболевало около 10 млн. человек. Вместе с тем в тот период эмпирически были выработаны:
• Принципы борьбы с эпидемиями (например, сжигание одежды больных, трупов умерших, изоляция пациентов).
• Открыты возбудители основных инфекционных болезней человека (сибирской язвы, дифтерии, столбняка и др.).
• Установлено, что патогенные для человека бактерии способны вырабатывать токсины, с непосредственным действием которых стали связывать развитие патологического процесса.
Аргументом в пользу важной роли бактериальных токсинов в развитии инфекционных болезней явилась высокая клиническая эффективность использования при них сывороток, что способствовало существенному снижению летальности от инфБ.
В последние годы, несмотря на предпринятые ранее эффективные меры, вновь стала расти заболеваемость туберкулёзом. Люди чаще стали повторно болеть теми инфекционными болезнями, после которых обычно вырабатывается стойкий иммунитет (корь, ветряная оспа и пр.). Нарастают сведения о важной роли некоторых возбудителей инфекционных болезней в формировании хронических заболеваний терапевтического (т.е. неинфекционного) характера.
Различают следующие виды инфекционного процесса.
• Сепсис — тяжёлая генерализованная форма инфекционного процесса, обусловленная размножением микроорганизмов в крови и нередко в других биологических жидкостях организма.
• Септикопиемия — инфекционный процесс, характеризующийся вторичным развитием гнойных очагов в различных тканях и органах у пациентов с сепсисом.
• Бактериемия, вирусемия — наличие в крови бактерий и/или вирусов без признаков их размножения. Является одним из этапов развития ряда инфП.
• Микстинфекция — инфекционный процесс, вызванный одновременно двумя возбудителями и более.
• Реинфекция — повторное (после выздоровления пациента) возникновение инфекционного процесса, вызванного тем же микроорганизмом.
• Суперинфекция — повторное инфицирование организма тем же возбудителем до периода выздоровления.
• Вторичная инфекция — инфекционный процесс, развивающийся на фоне уже имеющейся (первичной) инфекционой болезни, вызванной другим микроорганизмом.
Инфекционный процесс - сложный процесс взаимодействия возбудителя и макроорганизма в определённых условиях внешней и внутренней среды, включающий в себя развивающиеся патологические защитно-приспособительные и компенсаторные реакции.
Инфекционный процесс возникает при наличии трех компонентов:
- возбудитель,
- фактор передачи инфекции от заражённого организма к здоровому,
- восприимчивый макроорганизм (пациент).
Факторы, от которых зависит развитие инфекционного процесса:
- восприимчивость макроорганизма (реакция организма на внедрение возбудителя инфекции, развитие заболевания, или бакносительство);
- инвазивность возбудителя инфекции (способность микроорганизма проникать в ткани и органы макроорганизма и распространяться в них);
- доза возбудителя;
- патогенность возбудителя (способность микроорганизма в естественных условиях вызывать инфекционные заболевания);
- вирулентность возбудителя (степень патогенности данного микроорганизма при стандартных условиях естественного или искусственного заражения).
Восприимчивость хозяина к возбудителю зависит от многих факторов:
1 - неблагоприятная окружающая среда;
2 - нарушения иммунного статуса;
3 - наличие длительных хронических заболеваний;
4 - изменение нормальной микрофлоры кишечника (приём антибиотиков, стероидных гормонов);
5 - химио- и лучевая терапия;
6 - возраст;
7 - недостаточность/неполноценность питания;
8 - обширность травматических поражений, высокая степень ожогов и отморожений;
9 - низкая гигиеническая культура населения.
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 1540; Нарушение авторских прав?; Мы поможем в написании вашей работы!