КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Эксплуатация испытательных стендов ракетно-космических систем / Галеев А.Г., Золотов А.А., Перминов А.Н., Родченко В.В. Монография. Изд-во МАИ. – 2007
|
|
|
|
Экологические проблемы и риски воздействий ракетно-космической техники на окружающую природную среду. Справочное пособие / Под общей ред. В.В. Адушкина, С.И. Козлова и А.В. Петрова. – М.: Изд-во “Анкил”, 2004.
Галеев А. Г. Экологическая безопасность при испытаниях и отработке ракетных двигателей. Учебное пособие. М.: Изд-во МАИ, 2006, 92 с.
Основная литература
Оказание первой помощи…………………………………………………………..89
Доктор технических наук, профессор А. Г. Галеев
Гомеостаз— способность биологических систем — организма, популяции и экосистем — противостоять изменениям и сохранять равновесие. Исходя из кибернетической природы экосистем — гомеостатический механизм — это обратная связь.
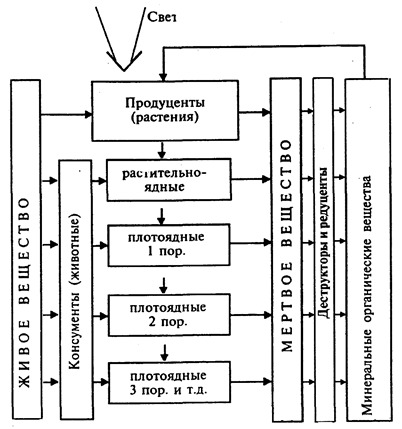
Рис.
Трофическая (функциональная) структура экосистемы (цепь питания) и круговорот вещества в ней
Следующий тип связей носит название форических (лат. форас -наружу, вон). Они возникают в том случае, если одни организмы участвуют в распространении других или их зачатков (семян, плодов, спор). Животными это распространение может осуществляться как на наружных покровах, так и в пищеварительном тракте.
Выделяют также тип связей, которые носят название фабрических (лат. фабрикатио - изготовление). Для них характерно использование одними организмами других или продуктов их жизнедеятельности, частей (например, растений, перьевого покрова, шерсти, пуха) для постройки гнезд, убежищ и т. п.
Взаимоотношения организмов. Данная классификация строится по принципу влияния, которое оказывают одни организмы на другие в процессе взаимных контактов. Эти взаимоотношения можно обозначить математическими значками «+», «–», «0» (положительно, отрицательно, нейтрально).
Если взаимоотношения обоим партнерам выгодны, они обозначаются значками (+,+) и носят название симбиоза или мутуализма. Степень этих связей различна. В ряде случаев организмы настолько тесно связаны, что функционируют как единый организм. Например, лишайники, представляющие симбиоз гриба и водоросли. Водоросль поставляет грибу продукты фотосинтеза, а гриб для водоросли является поставщиком минеральных веществ и, кроме того, субстратом, на котором она живет. В то же время сожительство грибов с корнями растений (микориза) носит хотя и взаимовыгодные, но не в такой степени тесные взаимоотношения. Тип взаимовыгодных отношений широко распространен. Сюда относятся и микроорганизмы, населяющие пищеварительный тракт животных, способствуя усвоению пищи; и, в ряде случаев, травоядные животные. Установлено, что исключение поедания трав животными может иметь следствием оскудение растительных сообществ, снижение ими продуктивности и устойчивости. Даже умеренное объедание листьев древесных растений насекомыми или их гусеницами может быть положительным не только для животных, но и для растений.
Взаимоотношения, которые положительны для одного вида и отрицательны для другого (+,-), характеризуются как хищничество и паразитизм. Хищник и паразит обычно приспосабливаются к использованию других организмов (их жертв и хозяев), а последние, в свою очередь, имеют адаптации, которые сохраняют им жизнь. Эти типы взаимоотношений обычно играют большую роль в регулировании численности организмов. Интенсивное размножение хищников и паразитов обычно имеет следствием уменьшение численности их жертв или хозяев.
В свою очередь, уменьшение численности жертв и хозяев подрывает кормовую базу хищников и паразитов, что ведет к сокращению их численности и т. д. В конечном счете имеет место обычно пульсирующая численность организмов, вступающих в такие типы взаимоотношений.
Хотя взаимоотношения типа хищничества и паразитизма сходны по результатам влияния на численность особей, они резко различаются по образу жизни и адаптациям. Во взаимоотношениях хищник-жертва оба организма постоянно совершенствуются: первый в плане успешности охоты, второй - в отношении самосохранения. И в том и в другом случае требуется быстрая реакция, высокая скорость передвижения, хорошее зрение, обоняние и т. п.
Во втором типе взаимоотношений у паразита адаптации идут по пути специализации структур на использование хозяина как источника пищи и «благоустроенного» местообитания. Результатом этого является упрощение многих органов (пищеварительный тракт, накожные покровы, органы передвижения, чувств и др.). Вместе с тем, поскольку жизнь паразита очень тесно связана с хозяином, он адаптирован на выживание во внешней среде после смерти хозяина. Достигается это за счет большого количества зачатков (семян, спор, цист и т. п.), обычно долго сохраняющихся в среде.
Адаптации хозяина направлены, как правило, на уменьшение вреда от паразита. Это проявляется в выработке активного иммунитета, заключении внутренних паразитов в различного вида капсулы (галлы, цецидии и т. п.).
В ряде случаев адаптации паразитов и хозяев приводят к их взаимовыгодным отношениям типа симбиоза. Есть основание полагать, что в большинстве случаев симбиоз (мутуализм) вырос из паразитизма.
Взаимоотношения, невыгодные обоим партнерам (-,-), носят название конкуренции. Последняя тем сильнее, чем ближе потребности организмов к фактору или условию, за которые они конкурируют. В этом отношении наиболее близки интересы организмов одного вида, и, следовательно, внутривидовая конкуренция рассматривается как более острая по сравнению с межвидовой. Однако данное положение противоречит тому факту, что практически все механизмы существования вида направлены на его выживание. Такое противоречие решается тем, что на внутривидовом уровне есть механизмы, которые позволяют снять остроту конкурентной борьбы, в том числе жертвуя частью особей (см. разд.V.2). Конкуренция и взаимоотношения типа хищник-жертва являются основными в совершенствовании видов, в то время как взаимоотношения типа мутуализма (симбиоза) способствуют оптимизации жизненных процессов, более полному освоению среды.
Менее распространенным типом взаимоотношений является комменсализм (франц. комменсал - сотрапезник) - отношения, положительные для одного и безразличные для другого партнера (+,0), его иногда делят на нахлебничество, когда один организм поедает остатки пищи со «стола» другого (крупного) организма (например, акулы и сопровождающие их мелкие рыбы; львы и гиены) и квартиранство, или синойкийю (греч. синойкос -сожительство), когда одни организмы используют другие как «квартиру», убежище. Например, молодь некоторых морских рыб прячется под зонтик из щупалец медуз, или некоторые насекомые живут в норах животных, гнездах птиц, используя их только для укрытия.
Не часто встречается также аменсализм (лат. аменс - безрассудный, безумный) - отрицательный для одного организма и безразличный для другого (-,0). Например, светолюбивое растение, попавшее под полог леса. Отношения, при которых организмы, занимая сходные местообитания, практически не оказывают влияния друг на друга, носят название нейтрализма (0,0). Например, белки и лоси в лесу. Сохранение разнообразия связей - важнейшее условие устойчивости экосистем.
Понятие экосистемы, ее состав, структура и свойства.
Состав экосистемы, как сказано ранее, представлен двумя группами компонентов: абиотическими - компонентами неживой природы и биотическими - компонентами живой природы, что общем виде в 1935году сформулировал Тенсли: { Экосистема } = [биоценоз] + [биотоп]. Позднее Сукачев (1947) это же понятие определил как биогеоценоз. Существенных расхождений в этих определениях нет, т.к. они полностью отражают синтез живой и неживой природы местообитаний в системах разного уровня и организации.
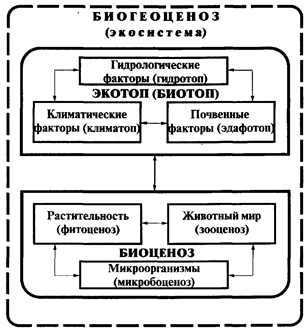
Рис 1 Схема биогеоценоза (экосистемы), по В.Н.Сукачеву
На рассмотрении этих составляющих окружающей нас природы остановимся более подробно.
Абиотические компоненты - это химические и физические элементы неживой природы:
- неорганические вещества и химические элементы, участвующие в обмене веществ между живой и неживой материей: диоксид углерода, вода, кислород, кальций, калий, натрий, железо, азот, фосфор, сера, хлор, и др.;
-органические вещества, связывающие абиотическую и биотическую части экосистем: углеводы, жиры, аминокислоты, белки, и др.;
-поток энергии;
-воздушная, водная или твердая среда обитания;
-климатический режим: солнечный свет, испарение, ветер, температура, влажность, осадки, водные течения и др.
Биотические компоненты состоят из трех функциональных групп организмов.
Первая группа организмов - продуценты или автотрофные организмы, т.е. "сами являющиеся пищей". В качестве питательного материала они используют простые неорганические вещества: воду, СО2, нитраты и фосфаты. В качестве энергетического материала продуценты используют солнечный свет либо энергию химических реакций. Они подразделяются на фото- и хемоавтотрофы.
Фотоавтотрофы используют в качестве источника энергии солнечный свет, а в качестве питательного материала - в основном углекислый газ и воду. К этой группе организмов относятся все зеленые растения и некоторые бактерии. В процессе жизнедеятельности они синтезируют на свету органические вещества - углеводы или сахара (СН2О)n,которыми питаются животные:
Фотосинтез: nСО2 + nН2О = (СН2О)n + nО2
Хемоавтотрофы используют энергию, выделяющуюся при химических реакциях. К этой группе принадлежат, например, нитрифицирующие бактерии, окисляющие аммиак до азотистой и затем азотной кислоты:
Хемосинтез: 2NH3 + 3O2 = 2HNO2 + 2H2O + Q
2HNO2 + O2 = 2HNO3 + Q
Химическая энергия (Q), выделенная при этих реакциях, используется бактериями для синтеза органических веществ.
Главная роль в создании органических веществ принадлежит зеленым растительным организмам. Роль хемосинтезирующих бактерий в этом процессе относительно невелика. Каждый год фотосинтезирующими организмами на Земле создается около 150 млрд. т органического вещества, аккумулирующего солнечную энергию.
Вторая группа организмов - консументы, или гетеротрофные организмы, т.е. "питающиеся другими". Они используют в качестве источника энергии и питательного материала готовое органическое вещество, осуществляя процесс разложения органических веществ. Их делят на фаготрофы и сапротрофы.
Фаготрофы питаются непосредственно растительными или животными организмами. К ним относятся в основном крупные животные - макроконсументы.
Сапротрофы используют для питания органические вещества мертвых остатков. К этой группе относятся как мелкие организмы (муравьи, черви и др.), так и крупные животные (гиены, шакалы, вороны и др.)
Третью группу организмов составляют редуценты или деструкторы, участвующие в последней стадии разрушения, т.е. в минерализации органических веществ, которые они восстанавливают до неорганических соединений (С, Н2, О2, N2, P и др.). Они очищают природную среду от отходов, возвращают вещества в кругооборот, превращая их в формы, доступные для продуцентов, таким образом возобновляя жизненный цикл.
К редуцентам относятся главным образом микроскопические организмы (бактерии, грибы и др.) - микроконсументы. Их выделяют в отдельную группу потому, что роль редуцентов в круговороте веществ чрезвычайно велика. Без них в биосфере накапливались бы груды органических остатков, иссякли бы запасы минеральных веществ, необходимых продуцентам, и жизнь в той форме, которую мы знаем, прекратилась бы.
Все названные группы организмов в любой экосистеме тесно взаимодействуют между собой, согласуя потоки вещества и энергии.
Трофические цепи. Закон Линдемана и качество энергии, экологические пирамиды.
Потоки энергии в пищевых цепях. В естественных природных экосистемах не существует отходов. Все организмы, живые или мертвые, являются пищей для других организмов. Перенос энергии пищи в процессе питания (основной источник жизнедеятельности в экосистемах) от ее источника через последовательный ряд живых организмов называется пищевой, или трофической цепью.
Трофические цепи – это путь однонаправленного потока солнечной энергии, поглощенной в процессе фотосинтеза, через живые организмы экосистемы в окружающую среду, где неиспользованная часть ее рассеивается в виде низкотемпературной тепловой энергии.
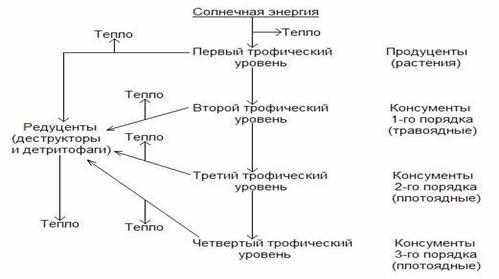
Трофические цепи делятся на два основных типа: пастбищные и детритные (лат. detrytys – продукт распада).
Пастбищная цепь тянется от зеленых растений к консументам: растительноядным животным и затем к плотоядным животным (хищникам).
Детритная цепь начинается с мертвого органического вещества – детрита, который разрушается детритофагами (лат. phagos – пожиратель), поедаемыми мелкими хищниками, и заканчивается работой редуцентов, минерализующих органические остатки.
Все организмы экосистем вовлечены в сложную сеть пищевых взаимоотношений. Пищевые цепи тесно переплетаются друг с другом, образуя пищевые, или трофические сети.
 Организмы, получающие энергию Солнца через одинаковое число ступеней, принадлежат к одному трофическому уровню. Так, зеленые растения – продуценты – занимают первый трофический уровень; травоядные животные – первичные консументы – второй; хищники – вторичные консументы – третий. Могут присутствовать хищники, поедающие первых хищников – третичные консументы, расположенные на четвертом уровне, и т.д. Но обычно наблюдается не более пяти уровней, так как на каждом уровне количество аккумулированной энергии резко падает (табл.).
Организмы, получающие энергию Солнца через одинаковое число ступеней, принадлежат к одному трофическому уровню. Так, зеленые растения – продуценты – занимают первый трофический уровень; травоядные животные – первичные консументы – второй; хищники – вторичные консументы – третий. Могут присутствовать хищники, поедающие первых хищников – третичные консументы, расположенные на четвертом уровне, и т.д. Но обычно наблюдается не более пяти уровней, так как на каждом уровне количество аккумулированной энергии резко падает (табл.).
Функциональные взаимосвязи, т.е. трофическую структуру, можно изобразить графически, в виде так называемых экологических пирамид. Основанием пирамиды служит уровень продуцентов, а последующие уровни питания образуют этажи и вершину пирамиды.
Известны три основных типа экологических пирамид: 1) пирамида чисел, отражающая численность организмов на каждом уровне; 2) пирамида биомассы, характеризующая массу живого вещества, — общий сухой вес, калорийность и т. д.; 3) пирамида продукции (или энергии), имеющая универсальный характер, показывающая изменение первичной продукции (или энергии) на последовательных трофических уровнях.
Простейшими из пирамид являются пирамиды численности, которые отражают количество организмов (отдельных особей) на каждом трофическом уровне. Для удобства анализа эти количества отображаются прямоугольниками, длина которых пропорциональна количеству организмов, обитающих в изучаемой экосистеме, либо логарифму этого количества. Часто пирамиды численности строят в расчёте на единицу площади (в наземных экосистемах) или объёма (в водных экосистемах).
В пирамидах численности дерево и колосок учитываются одинаково, несмотря на их различную массу. Поэтому более удобно использовать пирамиды биомассы, которые рассчитываются не по количеству особей на каждом трофическом уровне, а по их суммарной массе. Построение пирамид биомассы – более сложный и длительный процесс.
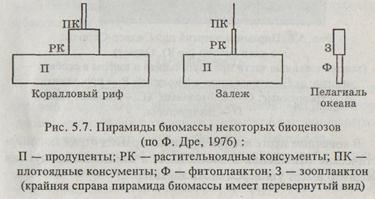
В наземных экосистемах действует следующее правило пирамиды биомасс: суммарная масса растений превышает массу всех травоядных, а их масса превышает всю биомассу хищников. Это правило соблюдается, и биомасса всей цепочки изменяется с изменениями величины чистой продукции, отношение годового прироста которой к биомассе экосистемы невелико и колеблется в лесах разных географических зон от 2 до 6%. И только в луговых растительных сообществах она может достигать 40—55%, а в отдельных случаях, в полупустынях — 70—75 %.
Наиболее фундаментальным и в определенном смысле идеальным способом отображения связей между организмами на разных трофических уровнях служит пирамида энергии, обладающая рядом преимуществ. Пирамида энергии наглядно иллюстрирует «правило десяти процентов»: На первом трофическом уровне в энергию пищи превращается лишь около 1% солнечного света. Вторичная продукция на каждом последующем трофическом уровне консументов составляет около 10% от предыдущей.
Эта закономерность носит название закона Линдемана, открытого им в 1942 г., или «правила 10%». Согласно этому закону в среднем только 10% от количества энергии, поступившей на трофический уровень, передается организмам, находящимся на последующем трофическом уровне. Так, количество энергии, которое доходит до третичных консументов, составляет около 10-4 энергии, поглощенной продуцентами. Это объясняет ограниченное число звеньев в пищевой цепи – редко до пяти-шести.
Пирамида энергии (продуктивности) любой экосистемы всегда имеет правильную форму, то есть подчиняется правилу: на каждом последующем уровне пищевой цепи продуктивность меньше, чем на предыдущем. Более того, суммарное количество вторичной продукции, образующейся на разных трофических уровнях, меньше первичной продукции данной экосистемы.

Продуктивность экосистем.
Продуктивность экосистем - это скорость, с которой живые организмы экосистемы производят полезную химическую энергию, заключенную в их биомассе. Продуктивность рассчитывается как количество энергии, аккумулированной организмами за единицу времени на единице площади [для наземных экосистем) или в единице объема (для водных экосистем).
Органическое вещество, производимое автотрофами, называется первичной продукцией. Скорость накопления энергии первичными продуцентами называется валовой первичной продуктивностью, а скорость накопления органических веществ – чистой первичной продуктивностью. ВПП примерно на 20 % выше, чем ЧПП, так как часть энергии растения тратят на дыхание. Всего растения усваивают около процента солнечной энергии, поглощённой ими.
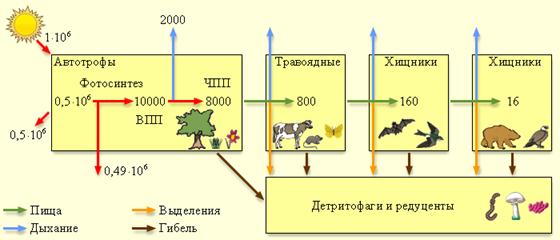
Рис. 2 Поток энергии через пастбищную пищевую цепь. Все цифры даны в кДж/м2·год
При поедании одних организмов другими вещество и пища переходят на следующий трофический уровень. Количество органического вещества, накопленного гетеротрофами, называется вторичной продукцией. Поскольку гетеротрофы дышат и выделяют непереваренные остатки, в каждом звене часть энергии теряется. Это накладывает существенное ограничение на длину пищевых цепей; количество звеньев в них редко бывает больше 6. Отметим, что эффективность переноса энергии от одних организмов к другим значительно выше, чем эффективность производства первичной продукции. Средняя эффективность переноса энергии от растения к животному составляет около 10 %, а от животного к животному – 20 %. Обычно растительная пища энергетически менее ценна, так как в ней содержится большое количество целлюлозы и древесины, не перевариваемых большинством животных.
Изучение продуктивности экосистем важно для их рационального использования. Эффективность экосистем может быть повышена за счёт повышения урожайности, уменьшения помех со стороны других организмов (например, сорняков по отношению к сельскохозяйственным культурам), использования культур, более приспобленных к условиям данной экосистемы. По отношению к животным необходимо знать максимальный уровень добычи (то есть количество особей, которые можно изъять из популяции за определённый промежуток времени без ущерба для её дальнейшей продуктивности)
Население Земли составляет около 6,0 млрд человек, каждому требуется ежегодно почти 1 млн ккал пищи, т.е. человечеству необходимо только на питание 6•1015 ккал энергии. В мире ежегодно собирается около 7·1015 ккал первичного органического вещества, но из-за неравномерного распределения, потерь и низкого качества части урожая этого количества энергии оказывается недостаточно. Кроме того, человек использует первичную продукцию не только как пищу, но и в виде волокон (хлопок, лен), и в качестве топлива (древесина и др.). В некоторых странах деревья сжигают гораздо быстрее, чем они могут расти, и леса превращаются в пустыни.
Описание потоков энергии является фундаментом экологического анализа для прогнозирования выхода полезных для человека продуктов.
Первичная продукция агроценозов и природных сообществ - основной источник пищи для человечества. Важна и вторичная продукция, так как животные белки включают ряд незаменимых аминокислот, которых нет в растительной пище. Пользуясь расчетами продуктивности экосистем, можно регулировать в них круговорот веществ, добиваясь выхода выгодной для человека продукции. Но необходимо хорошо представлять допустимые пределы изъятия растительной и животной биомассы (до 50÷60%), чтобы не разрушить экосистемы.
Экологические сукцессии.
 Закономерное развитие (изменение) экосистем во времени называется экологической сукцессией. Наблюдения показывают, что в результате естественных (движения ледников, пожаров, наводнений, землетрясений, извержений вулканов и др.) или антропогенных (пожаров, вырубки леса, добычи ископаемых, создания водохранилищ, загрязнения и т.д.) нарушений биоценозы и даже почва в экосистемах частично или полностью могут исчезать. После таких кардинальных изменений лица ландшафтов или агроценозов, смытые или эродированные почвы, или выжженный лес постепенно завоевываются многолетними дикими травами, затем кустарниками и, в конце концов, деревьями. Такой закономерный процесс, при котором биоценоз экосистемы с течением времени последовательно замещается серией других биоценозов, и является экологической сукцессией.
Закономерное развитие (изменение) экосистем во времени называется экологической сукцессией. Наблюдения показывают, что в результате естественных (движения ледников, пожаров, наводнений, землетрясений, извержений вулканов и др.) или антропогенных (пожаров, вырубки леса, добычи ископаемых, создания водохранилищ, загрязнения и т.д.) нарушений биоценозы и даже почва в экосистемах частично или полностью могут исчезать. После таких кардинальных изменений лица ландшафтов или агроценозов, смытые или эродированные почвы, или выжженный лес постепенно завоевываются многолетними дикими травами, затем кустарниками и, в конце концов, деревьями. Такой закономерный процесс, при котором биоценоз экосистемы с течением времени последовательно замещается серией других биоценозов, и является экологической сукцессией.
Выделяют два типа сукцессий: первичные и вторичные.
Первичная сукцессия начинается на участке, прежде не занятом живыми организмами и лишенном растительности. Такими участками могут быть скалы, глины после прохождения селя, остывшая вулканическая лава, районы открытой добычи полезных ископаемых. Вначале участки заселяют и формируют на них почвы несколько неприхотливых видов-пионеров. Например, поселения лишайников на камнях. Под действием их выделений каменистый субстрат постепенно превращается в подобие почвы, где поселяются кустистые лишайники, зеленые мхи, травы, кустарники и т.д.
В водных экосистемах эти виды постепенно образуют донные отложения. В итоге, виды-пионеры изменяют условия настолько, что участок становится более пригодным для новых групп растений и животных.
Вторичная сукцессия возникает там, где новое сообщество развивается на месте ранее существовавшего и где сохранились почвы или донные отложения. Такими территориями могут быть заброшенные сельхозугодья и посёлки, погребённые почвы, сведенные леса, загрязненные водоемы, затопленные при строительстве водохранилищ земли и т.д.
Скорость сукцессий различна. Для первичных сукцессий требуются сотни и тысячи лет. Вторичные протекают быстрее. Для восстановления растительной биомассы на месте вырубки, лесного пожара или покинутого сельскохозяйственного участка требуется от 20-50 до 250 лет.
Сукцессии обычно начинаются в незрелых несбалансированных сообществах, у которых скорости продукции органического вещества (П) либо больше, либо меньше скорости дыхания (Д). Сообщество стремится к более стабильному, зрелому состоянию, где П=Д. Сукцессия, начинающаяся при П>Д, называется автотрофной, а при П<Д - гетеротрофной. Отношение П/Д является функциональным показателем зрелости экосистем.
Состояние стабилизированной экосистемы называется климаксом.
Климаксныесистемы образуют сложную сеть взаимоотношений, поддерживающих их стабильное состояние. Теоретически такое состояние должно быть постоянным во времени и существовать до тех пор его не нарушат сильные внешние возмущения. Чем больше отношение П/Д отклоняется от 1, тем менее зрелой и устойчивой является экосистема. В климаксных сообществах это отношение приближается к 1.
А вот смены фауны и флоры в истории Земли похожи на медленно протекающие экологические сукцессии. Они тесно связаны с геологическими и климатическими изменениями и видообразованием. Такие процессы протекают на протяжении миллионов лет и называются эволюцией.
Эволюция экосистем, в отличие от сукцессий представляет собой уже длительные процессы их исторического развития, которые необратимы и ацикличны.
Саморегуляция и стабильность экосистем, гомеостаз. Устойчивость экосистем.
Относительно стабильное соотношение скоростей автотрофных и гетеротрофных процессов на Земле существует благодаря способности экосистем и биосферы к саморегуляции (гомеостазу).
Для управления экосистемами не требуется регуляция извне — это саморегулирующаяся система. Саморегуляция (управление) основана на обратных связях, когда часть сигналов с выхода из системы вновь поступает на вход, регулируя состояние системы на выходе. Саморегулирующий гомеостаз на экосистемном уровне обеспечен множеством управляющих механизмов. Один из них — субсистема «хищник—жертва».
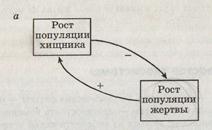
Обратные информационные связи необходимы для сохранения равновесия в экосистемах и бывают положительными и отрицательными.
Положительная обратная связь «усиливает отклонение», например, увеличивает чрезмерно популяцию жертвы. Отрицательная обратная связь «уменьшает отклонение», например, ограничивает рост популяции жертвы за счет увеличения численности популяции хищников.
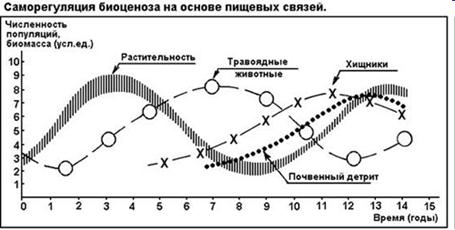
Наиболее устойчивы крупные экосистемы и самая стабильная из них – биосфера, а наиболее неустойчивы – молодые экосистемы. Это объясняется тем, что в больших экосистемах создается саморегулирующий гомеостаз за счет взаимодействия круговоротов веществ и потоков энергии.
Таким образом, гомеостаз - это способность популяции экосистемы самостоятельно поддерживать устойчивое динамическое равновесие при неразрушающих изменениях условий среды обитания с помощью обратных связей. Следовательно, экосистемы способны самостоятельно поддерживать относительную стабильность своего состояния.
Устойчивость экосистем
Стабильность в экологии означает свойство любой системы возвращаться в исходное состояние после того, как она была выведена из состояния равновесия. Стабильность определяется устойчивостью экосистем к внешним воздействиям. Выделяют два типа устойчивости: резистентную и упругую.
1.Резистентная устойчивость - это способность экосистемы сопротивляться нарушениям, поддерживая неизменными свою структуру и функции.
2.Упругая устойчивость - способность системы быстро восстанавливаться после нарушения структуры и функций.
Экосистеме трудно совмещать оба типа устойчивости: они связаны обратной связью, а иногда исключают друг друга. Например, калифорнийский лес из секвойи устойчив к пожарам (высокая резистентная устойчивость), но если он сгорит, то восстанавливается очень медленно или вовсе не восстанавливается (низкая упругая устойчивость). А заросли вереска очень легко выгорают (низкая резистентная устойчивость), но быстро восстанавливаются (высокая упругая устойчивость).
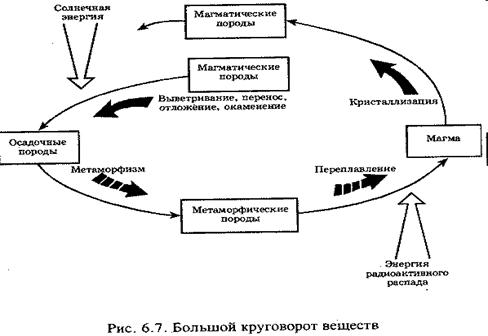
Круговороты веществ в биосфере. Большой (геологический) и малый (биологический) круговороты.
Глобальный круговорот веществ в биосфере.
Солнечная энергия обеспечивает на Земле два круговорота веществ: большой, или геологический (абиотический) и малый, или биологический (биотический).
Большой круговорот наиболее четко проявляется в циркуляции воздушных масс и воды. В основе большого (геологического) круговорота лежит процесс переноса веществ, в основном минеральных соединений, из одного места в другое в масштабе планеты.
Около 30% падающей на Землю солнечной энергии расходуется на перемещение воздуха, испарение воды, выветривание горных пород, растворение минералов и т.п. Движение воды и ветра, в свою очередь, приводит к эрозии почв и горных пород, транспорту, перераспределению, осаждению и накоплению механических и химических осадков на суше и в океане. В течение длительного времени образующиеся морские отложения могут возвращаться на поверхность суши, и процессы возобновляются. К этим циклам подключаются вулканическая деятельность, землетрясения и движение океанических плит в земной коре.
Круговорот воды, включающий ее переход из жидкого в газообразное и твердое состояния и обратно, - один из главных компонентов абиотической циркуляции веществ. В процессе гидрологического цикла происходят значительное перераспределение и существенная очистка планетарных запасов воды. При этом следует отметить, что наибольшей скоростью обновления обладают наиболее важные для существования живой среды суши – пресные воды. Период их оборота составляет в среднем около 11 суток.
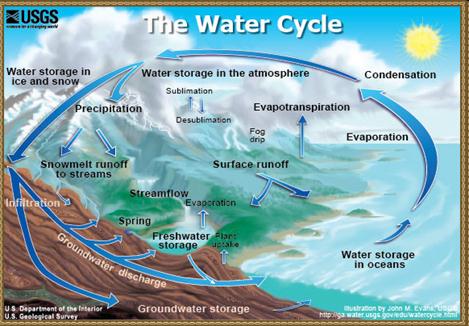
Малый круговорот веществ в биосфере (биогеохимический), в отличие от большого, совершается лишь в пределах биосферы.
Здесь действует биогеохимический круговорот, представляющий собой обмен макро- и микроэлементов и простых неорганических веществ (СО2 Н2О) с веществом атмосферы, гидросферы и литосферы. В основе малого круговорота веществ лежат процессы синтеза и разрушения органических соединений. Эти два процесса обеспечивают жизнь и составляют одну из главных ее особенностей.
Круговорот отдельных веществ В. И. Вернадский назвал биогеохимическими циклами. Суть цикла в следующем: химические элементы, поглощенные организмом, впоследствии его покидают, уходя в абиотическую среду, затем, через какое-то время, снова попадают в живой организм и т. д. Такие элементы называют биофилъными. Выделяют циклы С, О2, N2, P, S.
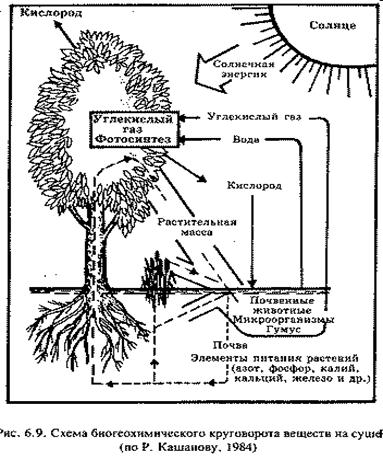
В отличие от геологического, биологический круговорот характеризуется ничтожным количеством энергии. На создание органического вещества, как уже упоминалось, затрачивается всего около 1% падающей на Землю лучистой энергии. Однако эта энергия, вовлеченная в биологический круговорот, совершает огромную работу по созиданию живого вещества. Чтобы жизнь продолжала существовать, химические элементы должны постоянно циркулировать из внешней среды в живые организмы и обратно, переходя из протоплазмы одних организмов в усвояемую форму для других.
В биогеохимических круговоротах следует различать две части, или как бы два среза: 1) резервный фонд — это огромная масса движущихся веществ, не связанных с организмами; 2) обменный фонд — значительно меньший, но весьма активный, обусловленный прямым обменом биогенным веществом между организмами и их непосредственным окружением. Если же рассматривать биосферу в целом, то в ней можно выделить: 1) круговорот газообразных веществ с резервным фондом в атмосфере и гидросфере (океан) и 2) осадочный цикл с резервным фондом в земной коре (в геологическом круговороте).
Круговорот углерода. В круговороте углерода, а точнее — наиболее подвижной его формы — СО2, четко прослеживается трофическая цепь: продуценты, улавливающие углерод из атмосферы при фотосинтезе, консументы — поглощающие углерод вместе с телами продуцентов и консументов низших порядков, редуцентов — возвращающих углерод вновь в круговорот. Скорость оборота СО2 составляет порядка 300 лет (полная его замена в атмосфере).
Главным резервуаром биологически связанного углерода являются леса, они содержат до 500 млрд т этого элемента, что составляет 2/3 его запаса в атмосфере. Вмешательство человека в круговорот этого элемента приводит к возрастанию содержания СО2 в атмосфере.
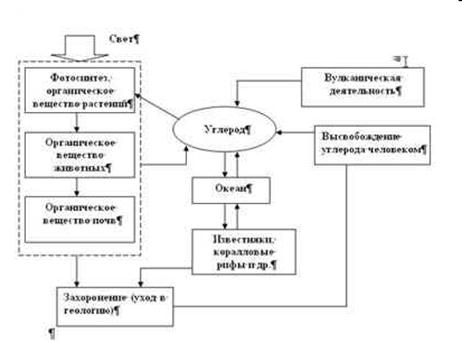
Углекислый газ из воздуха используется (усваивается) главным образом:
1) в процессе фотосинтеза СО2 + Н2О ® СН2О + О2;
2) в реакциях его с карбонатами океане СО2 + Н2О +СаСО3 ® Са(НСО3)2;
3) при выветривании горных пород Fе2S3 + 6СО2 + 6Н2О ®
2Fе(НСО3)3 + ЗН2S.
Поступление углекислого газа в атмосферу происходит в результате:
1) дыхания всех организмов;
2) минерализации органических веществ;
3) сжигания топлива для получения энергии и теплоты;
4) выделения при вулканических извержениях (незначительная часть - до 0,01%);
5) выделения по трещинам земной коры из осадочных пород (имеют также биогенное происхождение).
Круговорот кислорода. В значительной степени это антипод круговорота углекислого газа. Движение веществ одного происходит в направлении противоположном движению другого. Выработка O2 связана с реакцией фотосинтеза, тогда как выделение СО2 с дыханием. Общее потребление атмосферного кислорода и его возмещение первичными продуцентами происходит сравнительно быстро. Так, для полного обновления всего атмосферного кислорода требуется 2000 лет. Фотосинтез и дыхание в природных условиях, без учета деятельности человека, с большой точностью уравновешивают друг друга. В связи с этим накопления кислорода в атмосфере не происходит, и его содержание (20,946%) остается постоянным.

Роль кислорода в биосфере сложная, так как с ним в реакцию вступает большое количество органических и неорганических веществ. В результате возникает множество циклов от микроорганизмов до биосферы, происходящих между гидросферой, литосферой и атмосферой, между экосистемами данных сред, а также и внутри экосистем. Помимо обеспечения процессов жизнедеятельности кислород благодаря производному озону (O3), слой которого сформировался в стратосфере под действием ультрафиолетовой (УФ) радиации, осуществляет защиту живой природы от губительного жесткого УФ-излучения (λ < 280 нм).
Именно с появлением озона в атмосфере планеты (около 500÷600 млн. лет назад) связывается выход жизни из океана на сушу. До этого же момента большая часть кислорода, вырабатываемого в течение длительного геологического времени (около 3,0 млрд. лет), не оставалась в атмосфере, а фиксировалась литосферой в виде карбонатов, сульфатов, окислов железа и т.п. Это полностью исключало образование озона, а также способствовало его накоплению в литосфере, где масса кислорода в 15 раз превышает циркулирующий в биосфере O2 в виде газа или сульфатов, растворенных в континентальных и океанических водах.
Влияние антропогенной деятельности на снижение содержания кислорода в биосфере определяется теми же процессами, которые способствуют росту содержания оксидов углерода. В подтверждение этого достаточно привести лишь один пример – только автомобильным парком США сжигается кислорода в 2 раза больше, чем вырабатывается собственной природой США. Учитывая остальные потребности в кислороде, несложно понять размер «экологического долга» американцев только по этой «статье», а также назревающую остроту экологических проблем в промышленно развитых регионах планеты.
Круговорот азота. По общему объему составляет 1/65 от круговорота углерода и используется лишь 1/40 часть его резервного фонда. Крупнейшим резервуаром молекулярного азота N2 является атмосфера. Все живые организмы нуждаются в азоте, который используют в различных формах для образования белков и нуклеиновых кислот. Но лишь немногие микроорганизмы – азотофиксаторы могут использовать газообразный N2 из атмосферы. Однако, в процессе круговорота он преобразовывается в растворимые и усвояемые растениями ионы аммония NН4+ (аммонификация), нитрит- и нитрат-ионы NО2- и NО3- (нитрификация), что и способствует повышению его использования. Сложность круговорота азота заключается в том, что он включает газовую и минеральную фазы, а также абиотическую и биотическую части.
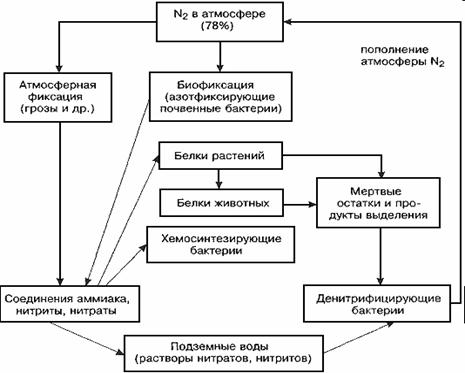
Антропогенное влияние на круговорот азота является разноплановым.
Антропогенная денитрификация (превращение избыточных нитритов и нитратов в газообразный N2) и стремление к сокращению производства нитратных удобрений соответствуют природным процессам поступления потока N2 в атмосферу, компенсируя тем самым его антропогенную фиксацию из атмосферы в сельском хозяйстве и промышленности.
В последнее время содержание N2 в атмосфере не менялось. Можно думать, что поступление его в атмосферу (денитрификация) и отток из атмосферы (азотфиксация) уравновешены, хотя фиксация слегка преобладает вследствие деятельности человека.
Следовательно, хотя человек и влияет как на потребление N2, так и на поступление его в атмосферу, эти потоки в целом сбалансированы и не меняют его концентрацию в воздухе, в отличие от потоков СО2
Круговорот фосфора. Круговорот фосфора в биосфере связан с процессами обмена веществ в растениях и животных. Этот важный и необходимый элемент протоплазмы, содержащийся в наземных растениях и водорослях (0,01÷0,1%) и у животных (от 0,1 % до нескольких процентов), циркулирует, постепенно переходя из органических соединений в фосфаты, которые снова могут использоваться растениями (рис.).
Однако фосфор в отличие от других биофильных элементов в процессе миграции не образует газовой формы. Резервуаром фосфора является не атмосфера, как у азота, а минеральная часть литосферы.
Ресурсы неорганического фосфора обеспечиваются ископаемыми изверженных (апатиты) или осадочных пород (фосфориты). Из материнской породы неорганический фосфор вовлекается в циркуляцию выщелачиванием и растворением в континентальных водах. Попадая в экосистемы суши, почву, фосфор поглощается растениями из водного раствора в виде неорганического фосфат-иона (РО43-) и включается в состав различных органических соединений, где он выступает в форме органического фосфата. По пищевым цепям фосфор переходит от растений к другим организмам экосистемы. Химически связанный фосфор попадает с остатками растений и животных в почву, где вновь подвергается воздействию микроорганизмов и превращается в минеральные ортофосфаты, а в дальнейшем происходит повторение цикла.
В водные экосистемы фосфор попадает благодаря поверхностному и грунтовому стоку. В пресных водоемах фосфаты вызывают бурный рост сине-зеленой растительности и деградацию водных экосистем. В соленых морских водах фосфор переходит в состав фитопланктона, служащего пищей другим организмам моря, в последующем накапливаясь в тканях морских животных, например, рыб. Отмершие остатки организмов приводят к накоплению фосфора на разных глубинах. Отсюда следует, что фосфор, попадая в водоемы тем или иным путем, насыщает, а нередко и перенасыщает их экосистемы.
Поэтому при рассмотрении круговорота фосфора в масштабе биосферы за сравнительно короткий период можно отметить, что он полностью не замкнут. Механизм возвращения фосфора из океанов на сушу в естественных условиях совершенно не способен компенсировать его встречный поток. В связи с тем, что запасы фосфора на Земле малы (содержание не превышает 1% в земной коре), то любые воздействия человека на биогеохимический круговорот фосфора несут опасность потери фосфора, что делает его менее замкнутым. П. Дювиньо (1967) подчеркивал, что «положение однажды окажется весьма угрожающим… фосфор — наиболее слабое звено в жизненной цепи, которая обеспечивает существование человека». А возрастающее антропогенное потребление фосфора существенно, если не угрожающе, усиливает опасную тенденцию быстрого истощения его запасов.
Круговорот серы. Существуют многочисленные газообразные соединения серы, такие, как сероводород H2S и сернистый ангидрид SO2., но преобладающая часть круговорота этого элемента имеет осадочную природу и происходит в почве и воде.
Основной источник серы, доступный живым организмам, — сульфаты (SO42-). Доступ неорганической серы в экосистеме облегчает хорошая растворимость многих сульфатов в воде. Растения, поглощая сульфаты, восстанавливают их и вырабатывают серосодержащие аминокислоты (метионин, цистеин, цистин), играющие важную роль в выработке протеинов.
В целом, по сравнению с азотом и фосфором сера реже является лимитирующим фактором для растений и животных. Вместе с тем круговорот серы относится к ключевым в общем процессе продукции и разложения биомассы. К примеру, при образовании в осадках сульфидов железа фосфор из нерастворимой формы переводится в растворимую и становится доступным для организмов. Это подтверждение того, как один круговорот регулируется другим.
Влияние антропогенной деятельности на вещественный баланс серы в биосфере незначительно и проявляется в основном в возникновении локальных опасностей, связанных с высокой токсичностью вырабатываемых серосодержащих веществ (газы, кислоты и соли).
Энергия в биосфере. Законы термодинамики и регуляторы энтропии в экосистемах.
Потоки энергии в биосфере.
Энергия – источник жизни, основа и средство управления всеми природными и общественными системами, а также одно из основных свойств материи – способность производить работу.
Все, что происходит внутри и вокруг нас, основано на работе, в процессе которой одни виды энергии переходят в другие согласно фундаментальным законам физики (термодинамики).
Превращение энергии Солнца в энергию пищи путем фотосинтеза, происходящего в зеленом листе, иллюстрирует действие двух законов термодинамики, которые справедливы и для любых других систем.
Первый закон термодинамики – закон сохранения энергии – гласит: энергия не создается и не исчезает, она превращается из одной формы в другую.
Закон подразумевает, что в результате превращений энергии никогда нельзя получить ее больше, чем затрачено: нельзя из ничего получить нечто. Однако на выходе из системы энергия преобразуется в иные формы.
Второй закон термодинамики утверждает: при любых превращениях энергия переходит в форму, наименее пригодную для использования и наиболее легко рассеивающуюся.
Действительно, солнечная энергия Qсолн, получаемая поверхностью зеленого листа, уравнивается рассеянной и концентрированной формами энергии в соответствии с первым законом термодинамики: Qсолн = qрасс + qконц. Лучистая энергия Солнца, попав на Землю, стремится превратиться в рассеянную тепловую. Доля световой энергии, преобразованной зелеными растениями в потенциальную энергию их биомассы, намного меньше поступившей (qконц < Qсолн). Большая часть энергии превращается в теплоту, покидающую затем и растения, и экосистему, и биосферу, в соответствии со вторым законом термодинамики (рис.) Пища, созданная в результате фотосинтезирующей деятельности зеленых растений, содержит потенциальную энергию химических связей, которая при потреблении ее животными организмами превращается в другие формы. Животные, поглощая пищу, также меньшую ее часть превращают в потенциальную химическую энергию синтезируемой ими протоплазмы, а большую часть переводят в теплоту, которая рассеивается в окружающем пространстве. Уровень рассеиваемой теплоты есть показатель энергетического беспорядка любых систем и данная неупорядоченность характеризуется энтропией – мерой количества связанной (рассеянной) потенциальной энергии, недоступной для использования. Однако, наряду с рассеянием в экосистемах происходит концентрация и качественное преобразование энергии в сторону ее более высокой эксергии, т.е. создаются структуры, обладающие низкой энтропией, что на первый взгляд противоречит второму закону термодинамики. Совместимость второго начала термодинамики со способностью живых систем создавать высокоорганизованные структуры и поддерживать в них порядок обосновал нобелевский лауреат И. Пригожин (1962, 1986, 1994). Он показал, что способность к самоорганизации может встречаться в системах, далеких от равновесных, но обладающих хорошо развитыми «диссипативными структурами» (рассеивающими структурами), откачивающими неупорядоченность. Упорядоченность природной экосистемы, т.е. структура биоценоза, поддерживается за счет дыхания всего сообщества организмов, которое постоянно «откачивает» из нее неупорядоченность, т.е. рассеивает лишнее тепло. Таким образом, биосфера, с уникальными функциями преобразования солнечной энергии в концентрированную энергию органических веществ в растениях и системой обратных связей является структурой энергетического упорядочивания. В противоположность этому, чем больше развита цивилизация, тем больше ее потребности в концентрированной энергии, запасаемой в природе. Три четверти энергии, потребляемой в современном мире, поступает от сжигания невозобновляемого ископаемого топлива: нефти, угля, природного газа. Следовательно, современная техносфера повышает энтропию окружающей среды, рассеивая упорядоченно концентрированную (в настоящем и прошлом) энергию. Итогом подобной «деятельности» может быть только разрушение биосферы и в гораздо больших масштабах, чем на любом предыдущем этапе человеческой истории.
Демэкология.Популяции - учебник " Экология" авторы Коробкин В.И., Передельский Л.В_.djvu - с. 86-103
Учение о биосфере - учебник " Экология" авторы Коробкин В.И., Передельский Л.В_.djvu - с. 155-230
СОДЕРЖАНИЕ
ВВЕДЕНИЕ.............................................................................................................................. 4
1. Особенности и области применения ракетных двигателей....... 6
1.1. Принципы устройства тепловых двигателей........................................................... 6
1.2. Общие особенности ЖРД........................................................................................... 10
1.3. Области применения ракетных двигателей............................................................ 11
2. Основные параметры ЖРД……………………………………………………………13
3. Требования к Двигательным установкам……………………………………15
4. Топлива тепловых двигателей……………………………………………………17
4.1. Факторы, определяющие требования к топливам.................................................. 17
4.2. Общие требования к топливам как термодинамической системе...................... 19
4.3. Топлива на основе воздуха в качестве окислителя (бензины, дизельное топливо, сжиженные газы, спирты)......................................................................................................................... 21
4.4. Жидкие ракетные топлива......................................................................................... 24
5. Оптимизация Двигательных установок в составе ЛА................... 31
6. Основные узлы и агрегаты ЖРД..................................................................... 33
7. Регулирование процессов и режимы работы ЖРД............................. 35
7.1.Основные задачи автоматики ЖРД и ее состав..................................................... 35
7.2. Системы управления конечными параметрами траектории движения ЛА (тяга и соотношение компонентов).............................................................................................................. 36
7.3. Система управления вектором тяги........................................................................ 41
7.4. Система наддува баков............................................................................................. 43
8. Запуск и останов ЖРД........................................................................................... 47
8.1. Основные требования к запуску и останову. Этапы запуска и останов….. 47
8.2. Системы раскрутки турбонасосного агрегата...................................................... 51
8.3. Система зажигания и воспламенения топлива.................................................... 55
9. Схемы ЖРД с насосной подачей топлива............................................... 60
9.1. Основные особенности схем.................................................................................... 60
9.2. Двигатели с насосной схемой подачи без дожигания генераторного газа...... 62
9.3. Двигатели с насосной схемой подачи с дожиганием генераторного газа....... 63
9.4. Удельные характеристики двигателей различных схем....................................... 65
10. ЭКОЛОГИЧЕСКИЕ ТРЕБОВАНИЯ ПРИ СОЗДАНИИ И ЭКСПЛУАТАЦИИ РАКЕТНО-КОСМИЧЕСКОЙ ТЕХНИКИ……………………………………………….……67
11. ПЕРСПЕКТИВЫ ПРИМЕНЕНИЯ ЭКОЛОГИЧЕСКИ ЧИСТЫХ КОМПОНЕНТОВ (МЕТАНА И ВОДОРОДА) В ЭНЕРГЕТИЧЕСКИХ И ТРАНСПОРТНЫХ СИСТЕМАХ..79
Приложение: 1.1. Охрана безопасности жизнедеятельности……………..87
2. Основные положения Федерального закона РФ «О промышленной безопасности…» …………………………………………………………………………....................90
Курс лекций «Экология испытаний ЛА» предназначен для студентов 4-го курса аэрокосмического факультета МАИ по специальности «Экология», содержит основы процессов и устройства ракетных двигателей и двигательных установок (ДУ), экологические требования к двигателям и ДУ, основы экспериментальной отработки (испытаний ЖРД и ДУ), основы устройства испытательнымхкомплексов и включает:
- лекции – 34 часа;
- практические занятия – 34 часов;
- экзамен.
2. Козлов А. А и др. Системы питания и управления жидкостных ракетных двигательных установок. Учебник / М.: Машиностроение, 1988, 352с.
3. Основы теории и расчета жидкостных ракетных двигателей: Учебник /
А. П. Васильев, В. М. Кудрявцев и др. / Под ред. В. М. Кудрявцева – 3-е изд. исп. и доп. М.: Высшая школа, 1983, 703с.
|
|
|
|
Дата добавления: 2013-12-13; Просмотров: 2027; Нарушение авторских прав?; Мы поможем в написании вашей работы!