КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Биоокисление аминокислот
|
|
|
|
Пищеварение белков
Белки начинают перевариваться в желудке. Белки, попадающие в желудок, стимулируют выделение гормона гастрина эпителиальными клетками желудка. Гастрин вызывает секрецию соляной кислоты обкладочными клетками желез слизистой желудка, а также пепсиногена главными клетками. Желудочный сок имеет рН от 1,5 до 2,5. Он действует как антисептик, убивая большинство бактерий. Кроме того, глобулярные белки подвергаются денатурации, их молекулы разворачиваются и вследствие этого внутренние пептидные связи полипептидных цепей становятся более доступными для ферментативного гидролиза. Пепсиноген превращается в активный пепсин путем автокатализа. В желудке пепсин гидролизует по карбоксильной группе те пептидные связи, которые образованы в основном ароматическими аминокислотами. В итоге из длинных полипептидных цепей образуется смесь более коротких пептидов.
Как только кислое содержимое желудка попадает в тонкий кишечник, в нем под влиянием низкого рН начинается секреция гормона секретина, поступающего в кровь. Этот гормон вызывает выделение из поджелудочной железы в тонкий кишечник бикарбоната, что приводит к нейтрализации рН желудочного сока. В тонком кишечнике переваривание белков продолжается. Поступление аминокислот в двенадцатиперстную кишку вызывает высвобождение гормона холецистокинина, который стимулирует секрецию нескольких ферментов поджелудочной железы с оптимумом в слабощелочной среде. Трипсиноген подвергается активированию с помощью фермента энтерокиназы, синтезируемой клетками кишечника, а также автокаталитически. Трипсин катализирует гидролиз полипептидов по карбоксильным группам остатков лизина и аргинина. Химотрипсиноген активируется трипсином. Химотрипсин гидролизует пептиды по карбоксильным группам ароматических аминокислот. Карбоксипептидаза синтезируется в поджелудочной железе в виде неактивного зимогена прокарбоксипептидазы и осуществляет последовательное отщепление С-концевых остатков. Аминопептидаза синтезируется клетками тонкого кишечника и осуществляет последовательное отщепление N-концевых остатков за исключением пролина. Дипептидаза гидролизует дипептиды.
В конце концов, свободные аминокислоты транспортируются через эпителиальные клетки, выстилающие тонкий кишечник, проникают в капилляры ворсинок и переносятся кровью в печень.
Катаболизм аминокислот
Белковый обмен в организме человека характеризуется высокой скоростью. Аминокислоты, поступающие в организм с пищей, расходуются на синтез белков, гормонов, нейромедиаторов, а неиспользованные подвергаются расщеплению. Аминокислоты в организме не запасаются. В процессе деградации азот аминокислот включается в структуру мочевины и выводится из организма с мочой, а их углеродный скелет либо превращается в липиды и углеводы, либо окисляется через центральные пути катаболизма.
Биологическая ценность пищевого белка зависит от качественного и количественного состава незаменимых аминокислот. Если белок содержит все незаменимые аминокислоты в необходимых соотношениях, то он является полноценным, если хотя бы одной незаменимой аминокислоты в белке нет, то биологическая ценность данного белка равна нулю.
Для оценки состояния обмена белков используют понятие азотистого баланса – разницы между количеством введенного с пищей азота и количеством выведенного в виде конечных продуктов азота в течение суток. Положительный азотистый баланс – количество выводимого из организма азота меньше количества азота, вводимого с пищей, - характерен для молодого растущего организма и свидетельствует о преобладании анаболических процессов. Отрицательный азотистый баланс – количество выводимого азота превышает количество поступающего азота – встречается при старении, тяжелых заболеваниях, голодании, когда преобладают катаболические процессы. Азотистое равновесие характерно для взрослого, здорового человека, находящегося на полноценной диете.
Катаболизм аминокислот начинается с отщепления у них аминогруппы в процессе дезаминирования или трансаминирования.
В процессе дезаминирования происходит отщепления от аминокислоты аминогруппы, которая превращается в аммиак. Наиболее важным является окислительное дезаминирование глутамата, осуществляемое глутаматдегидрогеназой, содержащей в качестве кофермента НАД+:
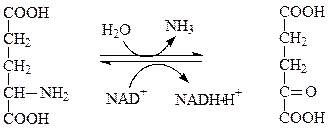
глутамат α-кетоглутарат
Глутаматдегидрогеназа – сложный аллостерический фермент, состоит из шести одинаковых субъединиц, активатор для нее – АДФ, ингибитор – ГТФ.
Аммиак может сохраняться, а затем использоваться для синтеза аминокислот. Реакция обратна предыдущей, но вместо НАД+ в ней участвует НАДФ+. Это обеспечивает независимую регуляцию этих двух реакций.
Трансаминирование – это реакция межмолекулярного переноса аминогруппы от аминокислоты на α-кетокислоту без промежуточного образования свободного аммиака. Механизм реакции является универсальным для всех живых организмов, протекает при участии ферментов аминотрансфераз, использующих пиридоксальфосфат в качестве кофермента.
Суммарное уравнение трансаминирования на примере трансаминирования аланина под действием аланинтрансаминазы (АЛТ):
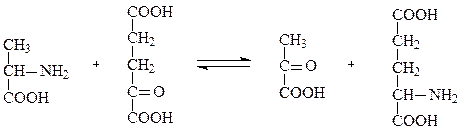
аланин α-кетоглутарат пируват глутамат
Реакции трансаминирования относятся к амфиболическим процессам, то есть выполняют как катаболическую функцию – отщепление от аминокислоты аминогруппы, так и анаболическую – синтез новой аминокислоты путем аминирования α-кетокислоты. Особую роль трансаминирование играет в процессах непрямого дезаминирования аминокислот. Непрямое дезаминирование протекает в две стадии. На первой стадии происходит трансаминирование аминокислоты с α-кетоглутаратом. При действии аминотрансферазы аминокислота превращается в кетокислоту, а α-кетоглутарат – в глутамат. На второй стадии происходит окислительное дезаминирование глутамата под действием глутаматдегидрогеназы и аминогруппа превращается в аммиак. Глутамат в этом процессе выполнил коллекторную функцию – собрал аминогруппы со всех аминокислот.
Важное значение при катаболизме аминокислот имеет декарбоксилирование с образованием биогенных аминов. Декарбоксилазы аминокислот также имеют пиридоксальфосфат в качестве кофермента.
При помощи фермента гистидиндекарбоксилазы из гистидина получается гистамин:
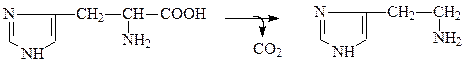
Глутаматдекарбоксилаза катализирует превращение глутамата в γ-аминомасляную кислоту:
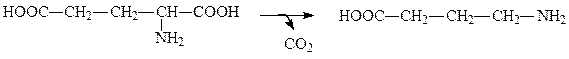
Двадцать различных катаболических путей, по которым идет расщепление аминокислот, в конечном счете сливаются и приводят всего лишь к шести продуктам, которые затем вступают в цикл Кребса и здесь окисляются полностью до СО2 и Н2О:
1) аланин, треонин, глицин, серин и цистеин превращаются в пируват;
2) фенилаланин, тирозин, лейцин, лизин и триптофан превращаются через ацетоацетил-СоА в ацетил-СоА;
3) аспартат и аспарагин превращаются в оксалоацетат;
4) фенилаланин и тирозин также могут превратиться в фумарат;
5) изолейцин, метионин и валин превращаются в сукцинил-СоА;
6) аргинин, гистидин, глутамин и пролин через глутамат превращаются в α-кетоглутарат.
Пути обезвреживания аммиака
Аммиак – крайне токсичное вещество, особо опасное для мозга. У большинства животных аммиак превращается сначала в нетоксичное соединение и лишь в таком виде переносится кровью от периферических тканей к печени или почкам.
Во многих тканях, включая мозг, NH3 взаимодействует с глутаматом в реакции, катализируемой глутаминсинтетазой, с образованием глутамина:

глутамат глутамил-5-фосфат глутамин
Глутамин – нейтральное, нетоксичное соединение, способное в отличие от глутамата проходить через клеточные мембраны. У большинства наземных животных глутамин доставляется кровью в печень, где он гидролизуется глутаминазой:
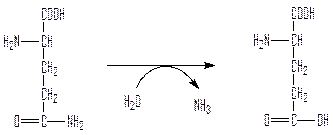
глутамин глутамат
В печени происходит затем превращение аммиака в мочевину. В крови здоровых людей содержание глутамина существенно превышает содержание других аминокислот.
Из мышц в печень аммиак переносится в виде аланина. В мышцах аммиак образуется при расщеплении аминокислот и при дезаминировании аденилата (АМФ) при напряженной работе. Существует глюкозо-аланиловый цикл:
Мышца Кровь Печень
Мышечный белок Глюкоза Глюкоза Глюкоза Мочевина
Аминокислоты NH3
NH3
Глутамат Пируват Пируват Глутамат
α-кетоглутарат Аланин Аланин Аланин α-кетоглутарат
Этот цикл выполняет две функции:
1) переносит аминогруппы из мышц в печень, где они превращаются в мочевину;
2) обеспечивает работающие мышцы глюкозой, поступающей с кровью из печени, где для ее образования используется углеродный скелет аланина.
В этом цикле аммиак превращается в аминогруппу глутамата в реакции, обратной окислительному дезаминированию, только глутаматдегидрогеназа использует НАДФН2 в качестве кофермента. Образовавшийся глутамат переносит затем свою α-аминогруппу на пируват, всегда имеющийся в достаточном количестве, поскольку это продукт гликолиза. Аланин – нетоксичная, нейтральная кислота, свободно выходящая из клеток.
Все животные делятся на три группы по способу выведения аммиака из организма:
А) Аммониотелические животные – большинство водных позвоночных, главным образом костные рыбы и личинки амфибий. У костных рыб аминный азот транспортируется кровью в виде глутамина, но через жабры он выводится в виде аммиака, так как в жабрах содержится глутаминаза. Костным рыбам не требуется сложной мочевой системы.
Б) Уреотелические животные – большинство наземных позвоночных, а также акулы. Так как свободный аммиак легко проникает сквозь мембраны и возможна его реабсорбция в почках и для выведения аммиака требуется одновременное выведение какого-либо аниона (хлорида или фосфата), то в ходе эволюции возникла способность выделять азот в виде мочевины – нейтрального, хорошо растворимого в воде и нетоксичного соединения. Однако на это затрачивается значительное количество АТФ.
В) Урикотелические животные – птицы, змеи, ящерицы. Для птиц очень важен их вес, а с мочевиной в мочу должны поступать и большие количества воды. Поэтому у птиц азот превращается в мочевую кислоту, которая относительно плохо растворима в воде, так что моча птиц – это полутвердая масса из кристаллов мочевой кислоты и очень небольшого количества воды. Биосинтез мочевой кислоты – это очень энергозатратный процесс. Мочевая кислота является также главным конечным продуктом обмена пуринов у приматов, птиц и рептилий.
Превращение аммиака в мочевину совершается в цикле мочевины.
Одна аминогруппа поступает в цикл мочевины в митохондриях при окислительном дезаминировании глутамата, вторую поставляет аспартат из цитозоля.
Карбомоилфосфатсинтетаза синтезирует высокоэнергетическое соединение карбамоилфосфат из аммиака, углекислого газа и АТФ:
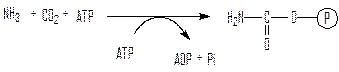
Орнитинкарбамоилтрансфераза переносит карбамоил на орнитин с образованием цитруллина:
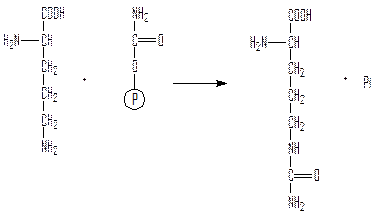
Аргининосукцинатсинтетаза синтезирует аргининосукцинат из цитруллина и аспартата:
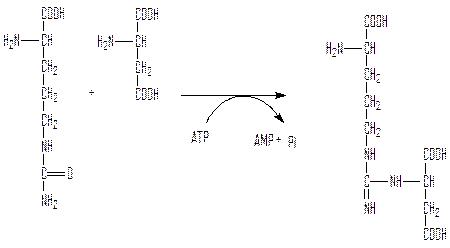
Аргининосукцинатлиаза расщепляет аргининосукцинат до аргинина и фумарата. Фумарат возвращается в пул промежуточных продуктов цикла Кребса.
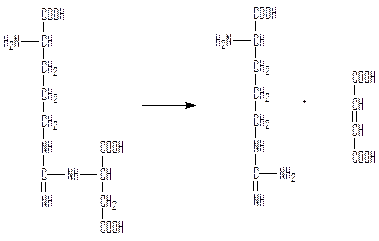
Аргинин может использоваться как аминокислота и тогда мочевина не образуется и цикл обрывается.
Аргиназа гидролизует аргинин с образованием мочевины и регенерацией орнитина, который может связать следующую молекулу карбамоила.
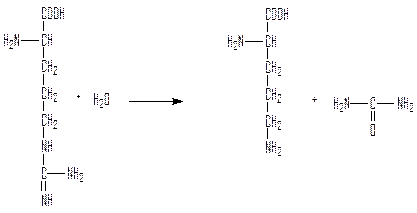
Суммарное уравнение цикла мочевины:
2NH3 + CO2 + 3АТФ + Н2О → NH2-CO-NH2 + 2АДФ + 2Pi + АМФ + PPi.
На синтез одной молекулы мочевины расходуется три высокоэнергетические фосфатные группы. У некоторых жвачных животных значительная часть мочевины поступает в рубец, где бактерии используют ее для синтеза аминокислот, которые затем всасываются и утилизируются хозяином.
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 1705; Нарушение авторских прав?; Мы поможем в написании вашей работы!