КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Види генетичних рекомбінацій
|
|
|
|
BOX
BOX
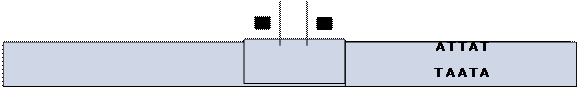
| Класи точкових мутацій | |
| Ген дикого | -А-Т-Т-Ц-Г- А -Ц-Т |
| типу | -Т-А-А-Г-Ц- Т -Г-Т |
| Транзиція | -А-Т-Т-Ц-Г- Г -Ц-Т |
| -Т-А-А-Г-Ц- Ц -Г-Т | |
| Трансверсія | -А-Т-Т-Ц-Г- Т -Ц-Т |
| -Т-А-А-Г-Ц- А -Г-Т | |
| Вставка | -А-Т-Т-Ц-Г- А - Г- Ц-Т |
| -Т-А-А-Г-Ц- Т - Ц- Г-Т | |
| Делеція | -А-Т-Т-Ц-Г---Ц-Т |
| -Т-А-А-Г-Ц---Г-Т | |
| Перестановка | -А-Т-Т-Ц-Г- Ц -Ц-Т |
| -Т-А-А-Г-Ц- Г -Г-А |
МУТАГЕНИ
Фізичні мутагени. Найкраще вивчена дія УФ-променів. Ультрафіолет викликає утворення тимінових димерів за рахунок виникнення додаткових зв’язків між 5 і 6 атомами піримідинових кілець (джерело помилок при реплікації).
Фотореактивація (світлова репарація)
Якщо безпосередньо після опромінювання ультрафіолетом подіяти видимим світлом (320-550 нм), то доля клітин, що вижили, буде в десятки разів більше. Світло активує фермент, який розчеплює димери тиміна і тільки їх, причому не має значення, ушкодження відбулися на одно- чи дволанцюговій ДНК.
Темнова реактивація (репарація)
 При темновій репарації вирізається сегмент ДНК (приблизно 30 п.н), добудовується з використанням неушкодженого ланцюга ДНК в якості матриці.
При темновій репарації вирізається сегмент ДНК (приблизно 30 п.н), добудовується з використанням неушкодженого ланцюга ДНК в якості матриці.


 -A-T- -A - T-
-A-T- -A - T-
-G-C- УФ -G - C-
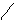
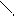 -T-A- (254нм) T A-
-T-A- (254нм) T A-
 -T-A- T A-
-T-A- T A-
-C-G- інактивація -C - G-
-G-C- -G - C-

 Видиме світло + Темрява + фермент
Видиме світло + Темрява + фермент
фермент
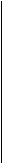
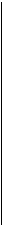


 -A T- - А - T- -A T-
-A T- - А - T- -A T-
-G C- -G C- -G C-



 -T A- -T A- -T A-
-T A- -T A- -T A-
-T A- -T A- -T A-
-C G- -C G- -C G-
 -G C- -G - C- -G C-
-G C- -G - C- -G C-
Фотореактивація Темнова реактивація
Дуже стійкий до впливу УФ-променів Deinococcus radiodurans (стара назва Micrococus radiodurans), які виявлені в уранових рудниках4 вони здатні витримувати 3-7 млн рад (для людини доза у 100 рад є смертельною).
Рентгенівські промені, γ-промені, протони, нейтрони теж можуть викликати мутації. Раніше згадувалось, що іонізуюче опромінення всіх видів частіше викликають одноланцюгові розриви в ДНК, на порядок менше розривів в обох ланцюгах.
Хімічні мутагени. Вони виявляють найрізноманітніший вплив на генетичний апарат клітин.
Аналоги азотистих основ дуже подібні за будовою до нормальних основ і включаються клітинами в ДНК. Найчастіше використовують бромураціл (аналог тиміну) та 2-амінопурин (аналог аденіну).
Кетоформа 5-бромуроцила, яка легко утворює пару з А, може таутомеризуватися в енольну форму, яка утворює пару переважно з Г, а не з А. Тоді при наступній реплікації основа Г обумовить включення Ц в комплементарний ланцюг. Аналогічно діє 2-амінопурин, який може «упізнаватись», як аденін і як гуанін.
|
|
|


 А—Т А—БУ БУ—Г Г—Ц
А—Т А—БУ БУ—Г Г—Ц
БУ
 Азотиста кислота викликає дезамінування амінопуринів (аденін) з утворенням гіпоксантина, який утворює пару з цитозином, а не з тиміном. В результаті відбувається неправильне спарювання і мутації типу АТ ГЦ.
Азотиста кислота викликає дезамінування амінопуринів (аденін) з утворенням гіпоксантина, який утворює пару з цитозином, а не з тиміном. В результаті відбувається неправильне спарювання і мутації типу АТ ГЦ.
Профлавін та інші акридинові барвники вбудовуються між сусідніми основами ДНК і збільшують відстань між ними (інтеркаляція). Такі просторові зміни при реплікації ДНК можуть призвести до серйозних порушень в ході зчитування інформації при синтезі білка (зсув рамки зчитування).
Алкілуючі агенти (етил-і метилметансульфонати, етиленамін, азотистий або сірчаний іприт) належать до найбільш ефективних мутагенів. Наприклад: етилметансульфонат, етилує (приєднує алкільні групи) до N-атому гуаніну. Утворюється 7-алкілгуанін, що відщеплюється від ланцюга, в результаті чого утворюється «пропуск». При наступному акті реплікації на цьому місці виявляється помилкова основа або делеція.
Біологічні мутагени — це фаг µ, міграційні генетичні елементи (IS (800-1400 пар основ), Tn — транспозони (2000 пар основ)). Вони інтегруються в різні ділянки хромосоми і викликають мутації типу інсерцій.
Для того, щоб виявити мутантів з певними цікавими для нас якостями існують різноманітні методи: середовища з додаваннями антибіотиків, метод відбитків, з використанням індикаторів.
РУХОМІ ГЕНИ
На початку 50-х років Барбара Мак-Клінтон, вивчаючи в Колд-Спрінг-Харборській лабораторії генетику кукурудзи, привернула увагу дослідників до незвичайної поведінки групи генів, які вона назвала «регуляторними елементами». Ці гени, ідентифіковані за їх властивостями подавляти експресію інших генів кукурудзи, які знаходилися з ними в близькому контакті, не мали фіксованого положення в хромосомах. Вони ніби рухались по геному рослини. Регуляторні елементи могли приєднуватись і відщеплюватись, при чому після їх відщеплення дуже часто відновлювались раніше «німі» гени. Виявилось, що гени, асоційовані з регуляторними елементами, стають не стабільними і часто мутували за нестабільності самих елементів.
Протягом багатьох років кукурудза зоставалась єдиною системою, в якій виявлялись такі рухомі генетичні елементи. Потім з’явились попередні дані, які свідчили про те, що деякі гени дрозофіли характеризуються високою частотою мутації і, напевно, також асоційовані з регуляторними елементами. Але більшість генетиків не звертали увагу на ці дані до тих пір, поки в кінці 60-х років не було доведено, що деякі плейотропні (тобто ті, що впливають на декілька різних функцій) мутації у E.coli обумовлені вбудовуванням в хромосому великих сегментів ДНК, названих інсерційними послідовностями (IS).
 IS1 IS1
 
    Ген термостабільного токсину І E.coli Ген термостабільного токсину І E.coli
Рис. 1. Інсерційна послідовність і транспозон, що нею генерується. IS1 містить ген ферменту транспозази, відповідальний за його переміщення. Цей ген фланкований повтореннями із 24 нуклеотидних пар (bp). В нижній частині малюнка зображений транспозон (Tn), що включає два таких IS-елемента, між якими знаходиться певний ген (в даному випадку ген термостійкого токсину, що викликає діарею). Важливо, що в багатьох незалежних актах інсерції брали участь одні і ті самі сегменти ДНК. Були знайдені 4 основних групи інсерційних послідовностей, названі IS1, IS2, IS3 і IS4. В хромосомі E.coli розкидані багаточисельні копії елементів IS1 і IS3. До пересування здатні не тільки індивідуальні IS-елементи; коли два таких елемента розташовані достатньо близько один від одного, вони можуть пересуватися як єдине ціле, захоплюючи лежачі між ними гени. Такі складні рухомі структури називаються транспозонами. З розвитком методів рекомбінантних ДНК з’явилась можливість клонувати IS-елементи всіх типів разом зі специфічними фланкуючими послідовностями. Порівняння послідовностей між IS-елементами і хромосомною ДНК показало, що вони можуть приєднуватись в багато ділянок хромосоми E.coli. Не менш важливим є те, що послідовності бактеріальної ДНК, фланкуючі IS-елементи зліва, завжди ідентичні послідовностям, що прилягають до правого кінця цього елемента. Така організація вказує на те, що сам акт інсерції, напевно, включає утворення в акцепторному сайті хромосоми розриву з липкими кінцями, між якими і приєднується донорний транспозон.
Переміщення транспозона полягає в утворенні нового дочірного транспозона Напевно, вираз «транспозон переміщається» не повністю точний, так як при появі транспозона в новій локалізації в первинному положенні він не зникає. Замість цього батьківський транспозон дає початок новій копії, яка приєднується в іншому місці бактеріальної хромосоми. Як це відбувається, до сих пір остаточно не відомо. Існує думка, що цей процес включає утворення розривів як у донорній, так і в акцепторній молекулі ДНК; потім донорна і акцепторна молекули з’єднуються з утворенням Х-подібних проміжних структур, в яких по дві сторони від транспозона знаходиться реплікативні вилки. В результаті реплікації утворюється два транспозона: один в первинному положенні, а другий - в акцепторній молекулі ДНК. Транспозиція закінчується сайт-специфічною рекомбінацією в межах транспозона, яка призводить до точного відновлення вихідної структури рухомого елемента і до утворення нового такого елемента, фланкованого знову утвореними прямими повторами. Таким чином, для транспозиції необхідна дія кількох сайт-специфічних нуклеаз. Одна із них вносить розрив з липкими кінцями на протилежних кінцях рухомих елементів, а інша-акцепторна область. Напевно, дві цих реакції каталізує один фермент - транспозаза; такі ферменти кодуються генами, розташованими в самих транспозонах. Мутаційний аналіз показав, що активність цього ферменту суворо необхідна для транспозиції. Встановлено, що синтез мРНК для транспозази регулюється білком-репресором, також кодуючим транспозоном. Зв’язуючись з відповідною операторною ділянкою в ДНК транспозона, цей репресор блокує синтез транспозази, в результаті чого утворення нової копії транспозона є дуже рідким явищем. Цікавим є те, що цей репресор також каталізує акт сайт-специфічної рекомбінації, яким закінчується транспозиція. Таким чином, при наявності відповідного сигналу репресор приймає участь у сайт-специфічній рекомбінації у ролі ферменту. Яким чином цей білок виконує дві дуже різних функцій, залишається незрозумілим. |


 24bp 720bp 24bp
24bp 720bp 24bp Tn1681
Tn1681

 IS1 552bp IS1
IS1 552bp IS1 Зигзагоподібний розрив в хромосомі
Зигзагоподібний розрив в хромосомі

 Транспозон
Транспозон



Транспозони мігрують по геному клітини і вбудовуються в різні ділянки даного генома та \ або здатні мігрувати з одного генома в інший. При гібридизації одноланцюгової ДНК, що містить IS - елемент з гомологічною ДНК без IS - елементів утворюються дволанцюгова ДНК з одноланцюговою «петлею» - це і є IS - елемент (найпростіший транспозон).
Tn містить 1 ген (для транспозази) або більше. Можуть вбудовуватися в плазміду. Переносять гени з плазміди в хромосому (є в дріжджах, маісі, дрозофілі, людині).
ГЕНЕТИЧНА РЕКОМБІНАЦІЯ
У еукаріот в процесі запліднення відбувається злиття двох гаплоїдних наборів генів, в результаті чого утворюються диплоїдна зигота. В ній проходить перекомбінація хромосом (кросинговер), і пізніше після редукційного поділу (мейозу) знов утворюються гаплоїдні гамети. Це статевий спосіб перетасовки генетичного матеріалу.
Бактерії завжди гаплоїдні. Але вони можуть передавати частину генетичного матеріалу з клітини-донора в клітину-реципієнта. Внаслідок цієї передачі утворюється так звана неповна зигота або мерозигота, яка містить частину генетичного матеріалу донора і всі гени реципієнта.
При наступному поділі ядра і клітини утворюються клітини з рекомбінантною хромосомою.
Рекомбінація - це процес в якому утворюється нова рекомбінантна хромосома, з генотипом, відмінним від обох батьків, шляхом комбінування генетичного матеріалу від двох організмів. Це процес обміну генетичним матеріалом, який належить різним особинам, в результаті чого виникають особини з рекомбінантним геномом. Нових генів в генофонді не з’являється — в цьому принципова відмінність рекомбінацій від мутацій.
Частину ДНК, що передається в клітину називають екзогенотою. Якщо ця ДНК стає стабільною частиною геному клітини-реципієнта, то вона стає ендогенотою. Ендогенотою така ДНК стає у двох випадках:
1. Коли вона інтегрується в хромосому.
2. Коли вона поступає в клітину у формі, недоступній клітинним ендонуклеазам. Тоді в неї немає необхідності інтегруватись в геном (наприклад у плазміди).
Незалежно від способу передачі ДНК екзогенота має лише 4 можливих долі в клітині:
1. Екзогенота може мати гомологічні ділянки з ендогенотою; інтегруватись в геном і призвести до утворення рекомбінантного геному.
2. Екзогенота іноді зберігається поза ендогенотою, реплікується утворюючи клон частково диплоїдних клітин.
3. Екзогенота може вижити, але не реплікуватись, тоді тільки 1 клітина стає частково диплоїдною.
4. Нуклеази клітин-господаря руйнуються екзогеноту, а процес називається рестрикція (вирізання).
1. Гомогогічна рекомбінація. Чужа ДНК рекомбінується з клітин ДНК шляхом обміну відповідними ділянками. Партнери повинні мати однакову нуклеотидну послідовність і мають бути максимально гомологічними.
2. Сайт-специфічна рекомбінація. Коротка дволанцюгова ДНК вбудовується в певному місці в довгу подвійну спіраль. Наприклад: інтеграція бактеріофага λ.
3. Негомологічна рекомбінація. Беруть участь негомологічні елменти ДНК. Це інтеграційна форма рекомбінації. На таку рекомбінацію здатні:
- вставочні (інсерційні) послідовні IS (ознак не кодують)
- траспозони Tn (містять гени стійкості до антибіотиків)
- бактеріофаг µ (він виконує роль і фагу, і гігантського транспозону)
У бактерій ДНК з клітини в клітину транспортується шляхом: кон’югації, трансдукції і та трансформації, а потім в клітині - реципієнті відбувається рекомбінація.
КОН’ЮГАЦІЯ
Кон’югація - це перенос генетичного матеріалу шляхом прямого контакту між клітинами. В 1946 р. Джошуа Ледерберг і Едвард Татум експериментально довели, що між бактеріями можливе свого роду спарювання.
Для дослідів були використані ауксотрофні мутанти E.coli, кожен з яких вимагав для росту 2 або 3 амінокислоти. Таким чином виключалася можливість простої реверсії (рис. 1).
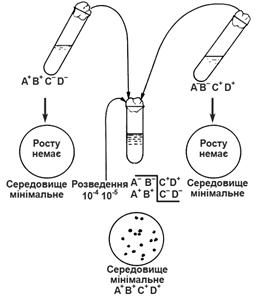
Рис. 1. Рекомбінація при кон’югації двох мутантів E.coli К-12
На чашках з’явились колонії з генотипом A+ B+ C+ D+.
Дослідження механізму кон’югації показали, що він починається з наближення клітин двох штамів і виникнення між ними кон’югаційного містка, в утворенні якого беруть участь F-пілі (fertility - плодючість).
Хромосома донора в певному місті розривається, потім реплікується і заново синтезований ланцюг 5’- кінцем проштовхується в середину клітини-реципієнта. Там відбувається рекомбінація між ДНК донора і реципієнта (рис. 2).
Кон’югація описана у бактерій родів Esherichia, Salmonella, Shigella, Pseudomonas, Nocardia. Здатність клітин бути донором пов’язана з наявністю у них статевого фактора F, яки й у разі кон’югації передається від однієї клітини до другої. Фактор F – це кільцева дволанцюгова ДНК (плазміда). Клітини, що не містять фактора F (F-), можуть функціонувати тільки як реципієнти. При кон’югації частота передачі фактора F наближається до 100%.
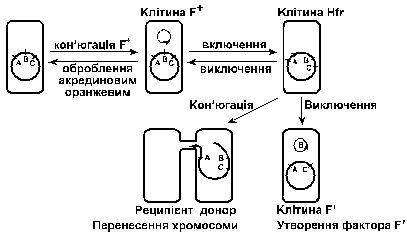
Рис. 2. Взаємовідносини між статевими типами E.coli (за Шлегелем, 1987)
У популяції F+ лише деякі клітини можуть бути донорами хромосомної ДНК. Це, зокрема, ті клітини, у яких фактор F інтегрувався в хромосому. Клітини-донори, що забезпечують високу частоту рекомбінації, називають Hfr (англ. higt frequency of recombination).
Як правило в клітину-реципієнт переноситься лише частина генетичного матеріалу клітини донора, в результаті чого утворюється неповна зигота — мерозигота, яка містить частину геному донора і повний геном клітини-реципієнта. Ділянки перенесеної від донора ДНК знаходять гомологічні ділянки в молекулі ДНК реципієнта, між якими проходить генетичний обмін. В результаті частина донорної ДНК вбудовується (інтегрується) в геном реципієнта, а відповідна частина реципієнтної ДНК з нього виключається. Ефективність інтеграції дуже висока: якщо донорний ген перенесений в клітину-реципієнт то ймовірність його включення складає приблизно 0,5.
За допомогою явища кон’югації можна виявити генетичну послідовність ДНК: перериваючи зв’язок між партнерами і вивчаючи, які гени були перенесені за певні проміжки часу, можна побудувати генетичну карту бактеріальної хромосоми. Перенос всієї хромосоми E. coli при температурі 370 C триває 100 хв.
ТРАНСДУКЦІЯ
Це явище описали у 1951 р. Леденберг і Циндер на прикладі фагу Р 22 у Salmonella typhimurium.
Трансдукція — передача ДНК від клітини-донора до клітини-реципієнта за участю бактеріофагів. Однак не всі фаги можуть здійснювати трансдукцію і не у всіх бактерій ДНК може бути перенесена таким шляхом. Відкрито у представників родів Esherichia, Salmonella, Shigella, Pseudomonas, Bacillus, Staphylococcus, Vibrio, Rhizobium.
Розрізняють два види трансдукції – загальну (неспецифічну) і специфічну. Неспецифічна трансдукція полягає у тому, що під час репродукції фага в клітині господаря, в капсиди замість фагової ДНК можуть включатися фрагменти ДНК бактеріальної хромосоми. Таким чином фаголізат містить суміш нормальних і дефектних фагів. Зараження клітин нормальним фагом веде до їх лізису. Зараження клітин бактеріальним дефектним фагом призводить до трансдукції (рис. 3).
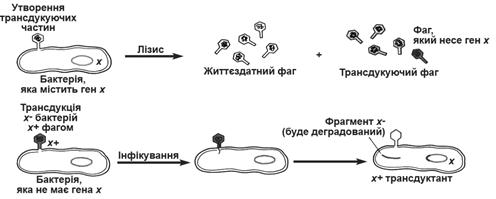
Рис. 3. Утворення фагових частин, які здійснюють загальну трансдукцію, і введення донорної ДНК в інфіковану клітину. Один із трансдукуючих фагів несе бактерійний ген х дикого типу, який при подальшому інфікуванні вводиться в бактерійну клітину, що несе мутантний ген х-.
При специфічній трансдукції бактеріофаг вбудовується чітко в певні ділянки бактеріальної хромосоми, наприклад., фаг λ встроюється між галактозним опероном і біоти новою ділянкою (gal і bio). Галактозний оперон відповідає за утилізацію галактози, біотинова ділянка – за синтез біотину - вітаміну H. Cирі яйця містять авідин - білок, який утворює з біотином нерозчинний комплекс.
ТРАНСФОРМАЦІЯ
Трансформація полягає в зміні властивостей одних бактеріальних клітин під впливом ДНК, яка виділена з інших бактеріальних клітин. Для трансформації не потрібний безпосередній контакт між клітинами. Гени можуть передаватися з клітини в клітину без всякого міжклітинного контакту і без переносників за допомогою вільної розчинної ДНК, що виділена з клітин-донорів.
Трансформація була вперше виявлена Фредом Гріффітом у 1928 р. при вивченні Streptococcus pneumonia (рис. 4). Він ввів мишам невелику кількість авірулентних безкапсульних клітин R-штаму (від англ. rough - гладкий) і вбиті нагріванням вірулентні капсульні клітини S-штаму (від англ. smooth - шершавий). Логіка експерименту полягала у наступному:
1. S-штам викликав загибель мишей;
2. R-штам не викликав загибелі;
3. S-штам, вбитий нагріванням не викликав загибелі;
4. Нагрітий S-штам + живий R-штам викликав загибель. З крові мишей, які загинули, були виділені вірулентні капсульні бактерії S-штама.
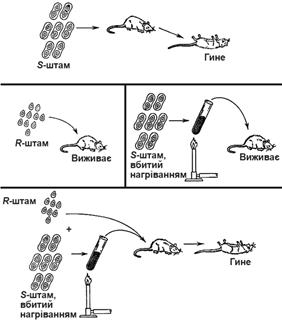
Рис. 4. Схема трансформації in vivo (дослід Ф.Гріффіта) (за Г. Стентом і Р. Келіндаром, 1981)
Це означало, що вбиті S-клітини передали здатність утворювати капсули R-клітинам за допомогою якогось фактора. Пізніше, в 1944 році, було встановлено, цим фактором є ДНК (Освальд Ейвері, Мак Леод, МкКарті).
Здатність ДНК проникати в клітину - реципієнту залежить як від природи самої ДНК, так і від фізіологічного стану самої клітини - реципієнта. Трансформуючою ДНК можуть бути тільки високомолекулярні дволанцюгові фрагменти. Проникати в бактеріальну клітину може ДНК, яка виділена з різних джерел (організмів), але включатися в геном — лише з певним ступенем гомологічності.
Після того, як екзогенний фрагмент ДНК, що проникає в клітину, знайшов гомологічний фрагмент ДНК клітини - реципієнта, між ними відбувається генетичний обмін аналогічно тому, як це має місце на останньому етапі кон’югації.
Проникний в клітину - реципієнта фрагмент спорідненої ДНК здатний викликати трансформацію з достатньо високою частотою – 0,2 - 0,3 (20%-30%).
Коли бактерії лізуються, то звільнюється певна кількість ДНК в оточуюче середовище. Ці фрагменти можуть бути відносно великими і містити декілька генів. Якщо такі фрагменти контактують з компетентними клітинами, то вони здатні проникати всередину клітини і здійснювати трансформацію. Це природній процес, який відбувається в ґрунті та на морі.
Генетична трансформація встановлена для родів Diplococcus, Bacillus, Rhizobium, Pseudomonas, Streptococcus. Цим способом передаються ознаки стійкості до різних отрут і прототрофність до окремих амінокислот. Трансформація здійснюється лише у певний період росту, коли клітини можуть поглинати молекули або фрагменти молекул ДНК. Цей період припадає здебільшого на середину експоненціальної фази росту. Період, коли клітина здатна поглинати ДНК, називається компетенцією. Компетентні клітини становлять приблизно 15% від всієї популяції.
СЕЛЕКЦІЯ МІКРООРГАНІЗМІВ
Дослідження геному мікроорганізмів, принципів його організації та функціонування має важливе значення для направленої селекції високопродуктивних штамів, що використовуються в промисловості.
У виробництві різних мікробних препаратів застосовують природні штами; штами, змінені в результаті мутацій; штами, одержані в методами генної і клітинної інженерії.
Природні штами використовуються здебільшого для виробництва біомаси, бактеріальних добрив і біоінсектицидів. Вони, як правило, не нагромаджують надлишку будь-якого метаболіту, оскільки у них наявні високоефективні регуляторні механізми. Тому у виробництвах, де цільовим продуктом є речовини, що виділяються клітинами у середовище, використовують генетично змінені штами.
Генетично змінені штами (штами, змінені в результаті мутацій) використовують у виробництвах, де одержують речовини, що виділяють клітини в середовище: продуценти антибіотиків, ферментів, органічних кислот, вітамінів, амінокислот.
Штами, які отримані методом генної інженерії. За допомогою методів передачі генетичної інформації в клітину-реципієнт створюються штами із заданими ознаками. Найчастіше використовують переносникі - вектори, плазміди, які вносять чужорідну ДНК і утворюють рекомбінантну (химерну) ДНК, а також протопласти, при злитті яких утворюються стабільні рекомбінанти. За допомогою генно-модифікованих штамів бактерій налагоджено промислове отримання інсуліну, соматотропіну, інтерферону тощо.
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 1369; Нарушение авторских прав?; Мы поможем в написании вашей работы!