КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Морфологія метафазних хромосом
|
|
|
|
Найбільш конденсовані хромосоми на стадії метафази мітозу, тому саме на цій стадії вивчають їх кількість і особливості будови. При диференційному забарвленні у хромосомах виділяються менш зафарбовані (еухроматинові) і більш зафарбовані (гетерохроматинові) ділянки. Вважають, що еухроматин являє собою менш конденсовані та більш активно транскрибовані ділянки хромосом.
Центромера (первинна перетяжка) поділяє хромосому на плечі. Центромери – це хромосомні структури, які відповідають за напрямок руху хромосом під час мітозу і мейозу. До функцій центромер належать когезія сестринських хроматид, утворення кінетохору, кон’югація гомологічних хромосом и залучення до контролю генетичної експресії. У більшості еукаріот центромери не містять визначеної послідовності ДНК. Зазвичай вона складається з великої кількості повторів ДНК (наприклад, сателітної ДНК), в якій послідовність усередині індивідуальних елементів, що повторюються, схожа, але не ідентична. У людини основна послідовність, що повторюється, називається α-сателітом, проте в цьому регіоні є кілька інших типів послідовностей. Встановлено, однак, що повторів α-сателіту недостатньо для утворення кінетохору і що відомі функціонуючі центромери, які не містять α-сателітної ДНК.
У визначенні місцеположення центромери у більшості організмів значну роль грають епігенетичні фактори. Дочірні хромосоми утворюють центромери в тих же місцях, що і материнська хромосома, незалежно від характеру послідовності, розташованої в центромерній ділянці. Передбачається, що повиний бути якийсь первинний спосіб визначення місцеположення центромери, навіть якщо згодом її місцеположення визначається епігенетичними механізмами.
ДНК центромери зазвичай представлена гетерохроматином, що істотно для її функціонування під час кон'югації та розходженні хроматид. У цьому хроматині людини (та щонайменш частини інших еукаріотів) нормальний гістон H3 заміщений центромеро-специфічним гістоном CENP-A. Вважається, що присутність CENP-A необхідна для збірки кинетохора на центромері і може грати роль в епігенетічному спадкуванні місцеположення центромери.
У нематоди Caenorhabditis elegans і деяких рослин (арабідопсис) хромосоми голоцентричні, тобто утворення кінетохора не локалізовано певною ділянкою, а відбувається дифузно по всій довжині хромосоми.
Саме в області центромери сполучені сестринські хроматиди в профазі і метафазі мітозу і гомологічні хромосоми в профазі і метафазі першого поділу мейозу. На центромерах же відбувається формування кінетохорів: білки, що зв'язуються з центромерою формують точку прикріплення для мікротрубочок веретена поділу в анафазі і телофазі мітозу і мейозу.
Кінетохор - білкова структура на хромосомі, до якої кріпляться волокна веретена поділу під час поділу клітини. Кінетохори відіграють найважливішу роль при сегрегації хромосом для подальшого поділу батьківської клітини на дві дочірні.
Кінетохори формуються на центромерах хромосом у еукаріотів. Кінетохори поділяють на дві області - внутрішню, міцно пов'язану з центромерною ДНК, і зовнішню, що взаємодіє з мікротрубочками веретена поділу.
На початку мітозу мікротрубочки, що швидко ростуть і тут же розпадаються, активно «обмацують» цитоплазму клітини в пошуках кінетохор. Зв'язки мікротрубочок, що кріпляться до одиничного кінетохору, називають «К-пучками» (англ. K-bundles). З ходом еволюції і ускладненням організмів, число мікротрубочок, що кріпляться до одиничного кінетохора, очевидно, зростала: у дріжджів виду Saccharomyces cerevisiae до кінетохору кріпиться одна мікротрубочка, у Schizosaccharomyces pombe - від 2 до 4, у мушки Drosophila melanogaster від 4 до 6, у ссавців - від 20 і більше.
Відхилення від нормального функціонування центромери ведуть до проблем у взаємному розташуванні хромосом в ядрі під час його поділу, і в результаті – до порушень процесу сегрегації хромосом (розподілу їх між дочірніми клітинами). Ці порушення приводять до анеуплоїдії, яка може мати тяжкі наслідки (наприклад, викликати синдром Дауна у людини, пов'язаний з анеуплоїдією (трисомією) по 21-ій хромосомі).
Кожна хромосома має два плеча, коротке (позначається p, від фр. pepite – «малий») і довге (q).
В залежності від місця розміщення центромери розрізняють метацентричні хромосоми (рівноплечі), субметацентричні (одне плече довше за друге), акроцентричні (одне із плеч дуже маленьке і коротке) і телоцентричні (хромосома закінчується центромерою) (рис.1). За сучасними уявленнями телоцентричних хромосом не існує, хромосоми повинні закінчуватися теломерами із деякою кількістю прицентромерного гетерохроматину. У 1938р. класики генетики Б.Мак-Клінток і Г. Меллер довели, що на кінцях хромосом є спеціальні структури, які назвали теломерами (телос-кінець, мерос-частина). Наприклад, акроцентричними є п’ять хромосом людини: 13, 14, 15, 21 і 22 (всі вони містять гени, що кодують рРНК). Телоцентричними є всі хромосоми миші, тоді як людина їх не має.
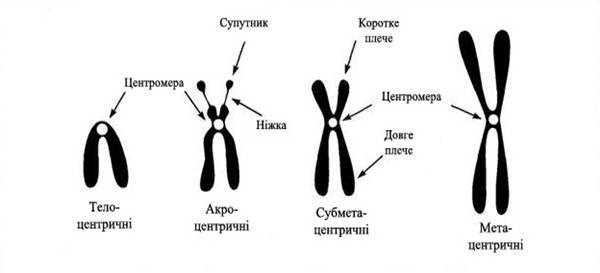
Рис.1. Типи метафазних хромосом
Теломери – специфічні кінцеві ділянки хромосом, містять паліндромні повтори нуклеотидних послідовностей, забезпечують індивідуальність хромосом (запобігають їх злиттю).
Вчені виявили, що при дії рентгенівським опроміненням стійкість проявляють лише теломери. Навпаки, позбавлені кінцевих ділянок, хромосоми починають зливатися, що веде до важких генетичних аномалій. Т.ч., теломери забезпечують індивідуальність хромосом. Теломери щільно упаковані (гетерохроматин) і малодоступні для ферментів (теломерази, метилаза, ендонуклеаз та ін)
Функції теломер.
1. Механічні: а) з'єднання кінців сестринських хроматид після S-фази, б) фіксація хромосом до ядерної мембрані, що забезпечує кон'югацію гомологів.
2. Стабілізаційні: а) запобігання від недореплікаціі генетично значущих відділів ДНК (теломери не транскрибируются), б) стабілізація решток розірваних хромосом. У хворих на α - таласемію в генах α - глобіну відбуваються розриви хромосоми 16д і до ушкодженого кінця додаються теломерні повтори (ТТАГГГ).
3. Вплив на експресію генів. Активність генів, розташованих поруч з теломерами, знижена. Це прояв сайленсінга - транскрипційного мовчання.
4. «Рахункова функція». Теломери виступають в якості часового пристрою, який відраховує кількість поділів клітини. Кожне ділення вкорочує теломери на 50-65 н.п. А всього їх довжина в клітинах ембріона людини становить 10-15 тисяч н.п.
Теломерна ДНК потрапила в поле зору біологів зовсім недавно. Перші об'єкти дослідження - одноклітинні найпростіші - війкова інфузорія (тетрахімена), яка містить кілька десятків тисяч дуже дрібних хромосом і, відповідно, безліч теломер в одній клітці (у вищих еукаріот менше 100 теломер на клітину).
Яке ж було здивування вчених, коли виявили, що теломерна ДНК людини відрізняється від такої у інфузорії всього лише однією літерою і утворює блоки 2 тимін - аденін - 3 гуанін (ТТАГГГ). Більше того, виявилося, що з ТТАГГГ - блоків побудовані теломери (Г - ланцюг) всіх ссавців, рептилій, амфібій, птахів і риб. Втім, дивуватися тут нема чому, тому що в теломерній ДНК не закодовано ніяких білків (вона не містить гени). У всіх організмів теломери виконують універсальні функції, мова про які йшла вище. Дуже важлива характеристика теломерної ДНК - їх довжина. У людини вона коливається від 2 до 20 тисяч пар основ, а у деяких видів мишей може досягати сотень тисяч п.н. Відомо, що поблизу теломер є спеціальні білки, що забезпечують їх роботу і беруть участь у побудові теломер. Доведено, що для нормального функціонування кожна лінійна ДНК повинна мати дві теломери: по одній теломері на кожному кінці хромосоми. У прокаріотів теломерів немає - їх ДНК замкнута в кільце.
Під час кожного циклу реплікації хромосоми вкорочуються на довжину РНК-праймера, що пов’язано із нездатністю ДНК-полімерази ІІІ будувати новий ланцюг де ново, а лише приєднувати нуклеотиди до готового ланцюга – праймера або затравки. Першим на проблему «кінцевий недореплікаціі ДНК» звернув увагу О.М. Оловніков в 1971 році. Він висловив гіпотезу про те, що втрата кінцевих послідовностей ДНК внаслідок їх недореплікаціі веде до старіння клітини. Передбачалося, що процес укорочення теломер і є той годинниковий механізм, який визначає реплікативний потенціал «смертної» клітини, і коли довжина теломер стає загрозливо короткою, цей механізм запобігає подальшому поділу клітини (ліміт Хейфліка). О.М.Оловніков припустив також, що в нестаріючих клітинах (а до них крім ракових відносяться зародкові, стовбурові та інші генеративні клітини) повинна існувати спеціалізована ферментативна система, яка контролює і підтримує довжину теломерной ДНК. Пізніше його припущення підтвердилось відкриттям ферменту теломерази. Теломерази найпростіших містять РНК довжиною в 150-200 нуклеотидних залишків (Н.О.), довжина теломеразної РНК людини - 450 н.з., в той час як теломераза дріжджів містить аномально довгу РНК (близько 1300 н.з.).
Крім центромери, хромосоми можуть мати вторинні перетяжки, а також супутники – ділянки хроматину, з’єднані з хромосомою тонкими тяжами.
|
|
|
|
|
Дата добавления: 2014-01-04; Просмотров: 2256; Нарушение авторских прав?; Мы поможем в написании вашей работы!