КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Репродуктивные органы цветковых растений
|
|
|
|
Покрытосеменные, или цветковые растения являются наиболее крупным отделом растительного царства, насчитывающим более 350 семейств, 13 тысяч родов и до 240 тысяч видов. В настоящее время покрытосеменные распространены по всему Земному шару – от тропиков до приполярных пустынь. Автотрофные цветковые растения – важнейший компонент биосферы, от которого зависят биосферные процессы обмена веществ и трансформации энергии, газовый состав атмосферы, климат, водный режим суши, характер процессов почвообразования. На суше цветковые часто доминируют в растительном покрове, только в немногих сообществах и типах растительности они уступают господство по общей массе другим группам растений – в хвойных лесах голосеменным, в некоторых типах тундр и на сфагновых болотах – мохообразным.
Покрытосеменные определяют самую возможность существования большинства наземных животных. Они формируют среду обитания для разнообразных консументов – потребителей первичного органического вещества. Различные группы гетеротрофов связаны с ними трофическими, консортивными связями. В процессе эволюции многие группы гетеротрофов смогли возникнуть только тогда, когда Землю завоевали покрытосеменные и образуемые ими сообщества. Для многих членистоногих и позвоночных характерна сопряженная с цветковыми эволюция. Человек как биологический вид тоже возник и эволюционировал после появления покрытосеменных, отличающихся высоким уровнем продуктивности и биохимическим разнообразием.
Покрытосеменные растения отличаются необычайным полиморфизмом, эволюционной пластичностью и могут существовать в разнообразных условиях обитания. Это единственная группа среди высших растений, представители которой сумели вторично освоить морскую среду, где стали доминировать на огромных акваториях.
Разнообразие размеров и жизненных форм цветковых растений, их приспособлений к конкретным условиям обитания велико. Представитель семейства рясковых – ряска малая (Lemna minor) не превышает 3-5 мм в длину и похожа на свободноплавающие слоевищные растения – мхи или водоросли (рис. 13.1).

Рис. 13.1. Ряска малая (Lemna minor) – миниатюрное цветковое растение.
С другой стороны, гигантские эвкалипты (виды рода Eucalyptus из семейства миртовые Myrtaceae) вырастают высотой до 100 м, длина побегов тропических лиан – ротанговых пальм измеряется сотнями метров. Среди покрытосеменных преобладают автотрофы, но встречаются полупаразиты, паразиты и сапротрофы, полностью утратившие хлорофилл и способность к фотосинтезу. Среди покрытосеменных имеются также эпифиты и эпифиллы (эпифиты, живущие на листьях других растений), деревья, кустарники, кустарнички, полукустарники и травы (многолетники и однолетники), плотоядные растения с их разнообразными приспособлениями для улавливания и переваривания мелких животных (рис. 13.2).

Рис. 13.2. Насекомоядное цветковое растение росянка круглолистная (Drosera rotundifolia).
Отмечены разнообразные приспособления видов цветковых растений к опылению строго определенными агентами, к распространению плодов и семян, к перенесению неблагоприятных климатических периодов. Эти особенности дают цветковым растениям возможность полностью реализовать свой эволюционно-адаптационный потенциал. Ни одна другая группа растений не смогла выработать такого разнообразия приспособлений к определенным факторам среды.
Существенным признаком покрытосеменных считают цветок – отсюда одно из названий отдела – Anthophyta (antos – цветок, phyton – растение). Происхождение цветка как особого органа – это одна из ключевых проблем морфологии и филогении покрытосеменных. Дискуссии по этому вопросу, начавшиеся еще в XVIII в., продолжаются и в настоящее время.
Типичный цветок определяют как видоизмененный побег, обеспечивающий функции спороношения, развития гаметофитов, половое размножение, образование семян в плоде. В истории ботаники отмечено, что в поисках архетипа цветка за первичный примитивный цветок различные ученые принимали разные структуры.
Сторонники эвантовой теории считают первичным крупный обоеполый цветок, опыляемый насекомыми, с большим и часто неопределенным числом свободных частей, и тогда цветки однополые, олигомерные, голые рассматриваются как результат более поздних этапов редукционной эволюции.
Сторонники псевдантовой теории полагают, что эволюционные трансформации выражались в постепенном усложнении структуры цветка от однополых, анемофильных, с небольшим числом частей к обоеполым, полимерным, энтомофильным, с хорошо развитым околоцветником.
Следует учитывать, что в процессе эволюционного возникновения покрытосеменные сначала приобрели комплекс важнейших общих признаков (в первую очередь – уникальные особенности жизненного цикла), а уже потом выработали разные цветки. При этом в каждой эволюционной ветви покрытосеменных были осуществлены различные морфогенетические решения.
Существенными частями цветка являются андроцей, состоящий из одной, нескольких или многих тычинок (микроспорофиллов) и гинецей, который состоит из одного, нескольких или многих плодолистиков, или карпелл (мегаспорофиллов).
Плодолистики (карпеллы) могут быть свободными, и тогда каждый из них, срастаясь краями, образует пестик (один или несколько пестиков в цветке – по числу плодолистиков). При срастании нескольких плодолистиков формируется ценокарпный гинецей, пестик в нем всегда один.
Важнейшая часть пестика – завязь. Наличие завязи (замкнутой полости), в которой развиваются семязачатки (в отличие от голосеменных, у которых семязачатки расположены на поверхности открытых мегаспорофиллов), обычно считают важнейшим признаком покрытосеменных, откуда и другое название отдела – Angiospermae (от греческого angeion – сосуд, вместилище, sperma – семя). Полагают, что завязь обеспечивает лучшую защиту семяпочек (семязачатков). В полости завязи семязачатки оказываются в условиях влажной камеры, оптимальных для их развития. Благодаря образованию завязи расширяются и обогащаются возможности распространения растений: из цветка формируется плод, при диссеминации расселяются не только отдельные семена, но и целые плоды, у которых возникают разнообразные приспособления к распространению широким кругом агентов.
У цветковых растений и некоторых голосеменных спермин (мужские неподвижные половые клетки) доставляются к яйцеклетке с помощью особой клетки – пыльцевой трубки, которая обеспечивает высокую точность попадания спермия в зародышевый мешок (то есть к женскому гаметофиту с яйцеклеткой). Семязачатки у покрытосеменных растений скрыты в завязи пестика, поэтому необходимо приспособление для улавливания пыльцы. Таким аппаратом у покрытосеменных служит рыльце на верхушке столбика пестика (стилодия). Наличие рыльца – важная особенность покрытосеменных растений.
Архегониальные растения (папоротникообразные, голосеменные) характеризуются тенденцией к возникновению разноспоровости, которая сопровождается редукцией полового поколения в жизненном цикле. Покрытосеменные, у которых редуцированы гаметангии, представляют собой завершающий этап на этом пути. Предельное сокращение процессов микро- и мегаспорогенеза и ускорение развития гаметофитов, связанное с их редукцией, были, вероятно, важными эволюционными факторами. Женский гаметофит цветковых растений представлен зародышевым мешком, состоящим всего из 7 клеток, а мужской гаметофит представлен пыльцевым зерном (пылинкой), в которой только 2 клетки – сифоногенная и спермагенная (дающая 2 спермия).
Уникальной особенностью покрытосеменных является двойное оплодотворение, в котором участвуют 4 клетки: два спермия, яйцеклетка и центральная клетка зародышевого мешка. В ходе этого процесса один из cnepмиев оплодотворяет яйцеклетку, образуя диплоидную зиготу, которая впоследствии развивается в зародыш семени. Второй спермий сливается с диплоидным ядром центральной клетки зародышевого мешка, образуя триплоидную зиготу. Эта зигота впоследствии дает начало триплоидному эндосперму семени. Общий для всех покрытосеменных уникальный процесс двойного оплодотворения указывает на их эволюционное единство и монофилетическое становление.
Одно из следствий двойного оплодотворения – формирование питательной ткани – полиплоидного эндосперма (в отличие от гаплоидного эндосперма голосеменных, представляющего остаток женского гаметофита). Биологическое значение полиплоидного состояния запасающей ткани связано с тем, что оно обеспечивает более благоприятные условия для развития зародыша и поэтому могло иметь важную эволюционную роль.
На тканевом уровне подавляющее большинство покрытосеменных характеризуется наличием в ксилеме сосудов, тогда как почти у всех голосеменных проводящие элементы древесины представлены только трахеидами. Только у немногих цветковых проводящая система ксилемы гомоксильная, бессосудистая, унаследовавшая примитивность от предковых голосеменных форм.
Паренхиматизация цветковых позволила выработать специальные запасающие ткани и травянистые жизненные формы. В отличие от древесных растений, у трав возможно сокращение онтогенеза и ускорение темпов эволюции; травянистые растения способны существовать в таких крайних условиях среды, в которых деревья и кустарники жить не могут.
Остается нерешенной проблема геологического возраста цветковых растений. Считалось, что покрытосеменные растения возникли раньше мелового периода – в триасе или палеозое (255-210 млн. лет назад). Предполагалось, что первоначально небольшая, эта группа существовала в таких условиях, которые не благоприятствовали фоссилизации их остатков, например, в горных районах или в пустынях. Позже, в меловой период (145-63 млн. лет назад), покрытосеменные дали мощную вспышку развития и сформировали свое современное разнообразие.
Этому взгляду противоречат объективные данные палеоботаники. Многочисленные указания на нахождение ископаемых остатков покрытосеменных в домеловых осадочных породах в настоящее время достоверно не доказаны. Имеются косвенные указания на возможность существования покрытосеменных в юре (210-145 млн. лет назад). Например, с этого времени известны некоторые формы насекомых, современные родственники которых настолько тесно связаны с цветковыми растениями, что допустить их независимое от последних существование трудно.
Древнейшие, достоверно определяемые ископаемые остатки цветковых растений в виде фоссилизованных листьев, цветков, плодов, древесины и пыльцевых зерен встречаются лишь с начала мелового периода (135 млн. лет назад). В этих отложениях они очень немногочисленны и фрагментарны. Еще недавно считалось, что к середине мелового периода (120 млн. лет назад) покрытосеменные появились в массе и выступали как «победители в борьбе за существование» с голосеменными и папоротникообразными. Сейчас палеоботаники отодвигают начало расцвета покрытосеменных на более позднее время – конец мелового периода (80-63 млн. лет назад).
Предполагают, что родиной цветковых могли быть тропические или субтропические регионы Земли. Исследователи считают, что это были районы тропической Юго-Восточной и Восточной Азии и Меланезия. В этом районе сосредоточены древние и считающиеся примитивными таксоны цветковых. Вероятно, что отсюда шла миграция древнейших цветковых в другие области Земли. Еще одна точка зрения заключается в признании центром происхождения покрытосеменны[ древнего континента Южного полушария – Гондваны. Считается, что цветковые могли возникнуть и расселиться по этому обширному континенту до начала раскола Гондваны на современные континенты Южного полушария.
К началу XX в. сложились две противостоящие друг другу системы взглядов на происхождение покрытосеменных от более примитивных предковых форм растений. Одна из них, связанная с именами австрийского ботаника Р. Веттштейна и классика немецкой ботаники А. Энглера, была основана на представлении о происхождении цветковых от эфедроподобных или гнетоподобных голосеменных предков. Р.Веттштейном была разработана оригинальная концепция происхождения цветка – так называемая псевдантовая гипотеза (от греческого «псевдантиум» – ложный цветок). Согласно этой гипотезе цветок представляет собой собрание мелких разнополых стробилов, претерпевших в процессе эволюции ряд преобразований: редукцию, сближение и срастание. На основе этой гипотезы наиболее архаичными следует считать семейства с тычиночными и пестичными невзрачными ветроопыляемыми цветками (ивовые, казуариновые).
Другая концепция восходит к построениям И.В.Гёте и А.П.Декандоля, то есть к концу XVIII – началу XIX веков. В эпоху филогенетической систематики это направление ознаменовалось появлением стробилярной (от греческого «стробилюс» – шишка хвойного)гипотезы, которую сформулировали Е.Арбер и Дж. Паркин. Другое название этой гипотезы – эвантиевая (от греческого «эвантиум» – настоящий цветок). Согласно стробилярной гипотезе наиболее вероятными предками покрытосеменных были мезозойские голосеменные растения – беннеттиты. С этих позиций цветок – это метаморфизированный (видоизмененный) укороченный спороносный побег, первоначально напоминавший шишку голосеменных. Мегаспорофиллы, несущие семязачатки, в процессе метаморфоза превратились в плодолистики (карпеллы), а несущие пыльники микроспорофиллы – в тычинки. Эти видоизменения многие исследователи связывают с приспособлением покрытосеменных к насекомоопылению (энтомофилии). Примитивные цветки имели значительные размеры и выпуклое цветоложе, на котором располагались многочисленные микро- и мегаспорофиллы. Микроспорофиллы эволюционировали в тычинки, а мегаспорофиллы – в карпеллы (плодолистики), которые образовали пестики. Цветки приобрели строение, сходное с тем, которое наблюдается у современных магнолиевых, лютиковых, кувшинковых. Поэтому перечисленные семейства обычно располагаются в основании системы покрытосеменных как наиболее древние и архаичные. Дальнейшая эволюция цветка в пределах покрытосеменных имела редукционный характер.
Обе эти гипотезы являются фолиарными (от латинского «фолиум» – лист), так как исходят из представлений, что цветки образовались из листостебельных спороносных побегов. Фолиарным гипотезам противопоставляются различные теломные (от греческого «телома» – конечные побеги) гипотезы (например, теория гонофилла Р. Мелвилла), широко распространенные в 60-70-е годы ХХ в. Согласно этим гипотезам, все части цветка могут быть выведены из теломов – осевых структур, свойственных ископаемым растения – риниевым.
Палеоботаника в последние десятилетия дала много нового материала о древнейших, ранне- и среднемеловых покрытосеменных. Эти данные позволяют сделать несколько заключений. Во-первых, уже самые ранние из известных нам цветковых отличаются большим полиморфизмом. Очень разнообразны пыльцевые зерна по форме, размерам, скульптуре поверхности, характеру поровости, числу и строению апертур. Они обнаруживают сходство с пыльцевыми зернами порядков и семейств современных растений. Листья меловых цветковых также весьма разнообразны и напоминают современные фикусы, протейные, дубы, акации, аралию, подорожник, известны и специализированные щитовидные листья.
Большим полиморфизмом отличаются и органы размножения – цветки и плоды. У меловых цветковых известны как одиночные и относительно крупные цветки, так и мелкие, собранные в разнообразно устроенные соцветия с простым или двойным околоцветником, с апокарпным или ценокарпным гинецеем, с верхней или нижней завязью, с разным числом тычинок и плодолистиков. Наряду со вскрывающимися плодами типа листовок обнаружены и не-вскрывающиеся плоды. Вместе с древесными формами в меловую эпоху существовали многочисленные травянистые покрытосеменные.
Во-вторых, если раньше часто утверждали, будто в меловую эпоху покрытосеменные были представлены современными таксонами, то сейчас большинство исследователей считают, что раннемеловые Angiospermae принадлежали к вымершим порядкам и семействам. Этот факт важен как доказательство того, покрытосеменные не оставались неизменными с момента зарождения.
В целом сейчас большинство специалистов считают, что предков Angiospermae нужно искать среди каких-то древних и малоспециализированных голосеменных растений. Не оставлена идея их генетической связи с беннеттитовыми. Допускают, что магнолиевые (как наиболее примитивные цветковые) лишь сохранили наибольшее число архаичных признаков, но сами, как и все прочие таксоны цветковых, произошли от какой-то вымершей, еще более древней гипотетической группы покрытосеменных.
В настоящем учебном пособии принята система цветковых растений, разработанная А.Л.Тахтаджяном в 1987 г. В данной системе древнейшей группой покрытосеменных считается порядок магнолиевых (Magnoliales), от предков которых, вероятно, произошли современные покрытосеменные. В этом смысле приведенная система монофилетична (рис. 13.3).
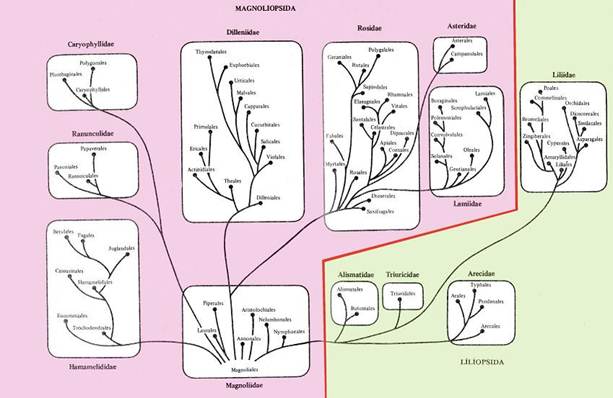
Рис. 13.3. Система покрытосеменных растений (по А.Л.Тахтаджяну).
Цветок – особый побег покрытосеменных растений, ответственный за формирование спор двух типов (микро- и мегаспор), развитие мужского и женского гаметофитов, образование гамет (яйцеклеток и спермиев), опыление, оплодотворение и формирование семян. Цветок в процессе своего развития превращается в плод с семенами. Во многих отношениях он уникален и столь характерен для этой группы, что покрытосеменные нередко называют цветковыми растениями. Исключительная роль цветка как особой морфологической структуры связана с тем, что в нем полностью совмещены все процессы бесполого и полового размножения. В цветках осуществляются микро- и мегаспорогенез, развитие гаметофитов, микро- и мегагаметогенез, опыление, оплодотворение и формирование зародышей нового поколения спорофитов. Завершается онтогенез цветка образованием плода с семенами. Особенности строения цветка обеспечивают осуществление всех этих функций с наименьшими затратами пластических веществ и энергии.
|
|
|
|
Дата добавления: 2014-01-05; Просмотров: 975; Нарушение авторских прав?; Мы поможем в написании вашей работы!