КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Характеристика спороутворювальних бактерій
|
|
|
|
ЕНДОСПОРИ ТА ІНШІ ФОРМИ СПОКОЮ У БАКТЕРІЙ.
До утворення спор здатна лише невелика група бактерій. Велике значення спор пов'язано з їх термостійкістю. У той час, як майже вся решта бактерій, а також вегетативні клітини споро-утворювальних видів гинуть при 80 °С (температура пастеризації) через 10 хв, терморезистентні спори витримують кип'ятіння впродовж кількох годин. Трудомістка та дорога техніка стерилізації (знезараження) розрахована на знищення спор. З іншого боку, терморезистентність спор є своєрідною можливістю вибіркового збагачення культур спороутворювальних форм. Ґрунт чи інший матеріал (мул, сіно, вода) прогрівають упродовж 10 хв при 80 °С. Така операція призводить до загибелі вегетативних клітин, і тільки терморезистентні спори залишаються життєздатними і проростають у відповідному поживному середовищі.
Спороутворювальні бактерії, за одним винятком, належать до паличкоподібних грампозитивних бактерій. Грамнегативними спороутворювальними бактеріями є представники роду Desulfotomaculum. Описано понад 10 родів бактерій, які утворюють ендоспори, наприклад, Bacillus, Clostrldium, Desulfotomaculum, Sporo lactobacillus, Oscillospora та ін. Більшість із цих бактерій є рухливими завдяки перитрихально розміщеним джгутикам. Бактерії, які належать до роду Bacillus, є строгими аеробами або факультативними анаеробами. Роди Clostridium і Desulfotomaculum об'єднують анаеробні бактерії, здатні до утворення спор. Клостридії одержують енергію за рахунок бродіння, види Desulfotomaculum — шляхом анаеробного дихання. SporolactobaciUus належить до молочнокислих бактерій. Sporosarcina має сферичні клітини, але за своїми фізіологічними ознаками належить до бацил.
Характерною особливістю спороутворювальних бактерій є низький вміст ГЦ (гуанін + цитозин) у ДНК клітин. Так, у клостридій ДНК містить від 22 до 27 % ГЦ.
Спори різних видів бактерій розрізняються за формою, розміром, положенням у клітині (рис. 2.10). За формою вони можуть бути круглими, овальними. Поверхня спор буває гладенькою, хвилястою, остеподібною та ін.
Спори можуть займати таке положення в клітині:
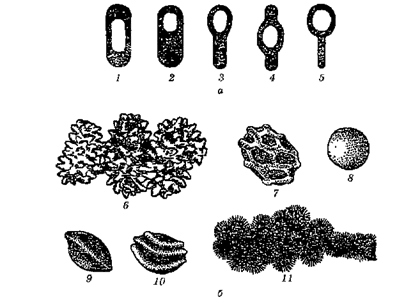
Рис.2.10. Розміщення спор у клітині (а) і поверхня спор (6):
1 — Bacillus megaterium; 2 — В. thuringiensis; 3 — Clostridium polymyxa; 4 — С. butyricum; 5 — С. tetani; 6 — остеподібна; 7 — з полігональними опуклостями; 8 — гладенька; 9 та 10 — хвиляста; 11 — остеподібна у конідієспор актиноміцетів
1) бацилярне, коли спора локалізується в клітині центрально, ексцентрально чи термінально, і при цьому клітина не змінює своєї форми. Такі клітини називають бацилами (Bacillus);
2) клостридіальне, коли при формуванні спори клітина змінює свою форму, набуваючи вигляду човника чи веретена. Такі клітини називають клостридіями (Clostridium, від грец. clos-ter — веретено);
3) плектридіальне, коли спора локалізується термінально, у місці її розміщення клітина розширюється і набуває форми ракетки. Клостридіальний і плектридіальний типи розміщення спор властиві в основному видам роду Clostridium і нерідко одночасно зустрічаються в культурі одного виду.
Виявлення ендоспор. При мікроскопічному дослідженні спори видимі завдяки своєму високому показникові заломлення світла — такому самому, як у знезводненого білка. Це вказує на те, що в спорах велика кількість багатого білком матеріалу сконцентрована в малому об'ємі. Спора містить майже всю суху речовину материнської клітини, але займає в 10 разів менший об'єм. У сумнівних випадках виявити в клітинах істинні ендоспори можна за допомогою спеціального фарбування. Якщо препарат бактерій, фіксований нагріванням, прокип'ятити з карболовим роз
чином фуксину, то спори міцно зв'язують барвник і не знебарвлюються навіть при обробці етанолом чи ЇМ оцтовою кислотою (в умовах, коли решта вмісту клітини стає безбарвною),
3.2. Спороутворення (споруляція)
Спори утворюються всередині бактеріальної клітини. Цей процес починається з накопичення білкового матеріалу, тому показник заломлення у місці утворення спори збільшується. При цьому відбувається споживання запасних речовин (полі-в-гідроксимасляної кислоти у аеробів і полісахаридів у анаеробів). Упродовж перших 5 год спороутворення значна частина білків материнської клітини розкладається. При цьому утворюється специфічна для спор речовина — дипіколінова кислота, яка не зустрічається у вегетативних клітинах. У ході синтезу дипіколінової кислоти відбувається поглинання іонів кальцію; очевидно, в зрілих спорах ця кислота міститься у вигляді хелату з кальцієм і може становити 10—15 % сухої речовини спор. Дипіколінова кислота міститься тільки в терморезистентних спорах.
Спороутворення є одним з найскладніших процесів диференціації бактеріальної клітини. Воно починається з особливого нерівномірного поділу клітини (рис.2.11).
У результаті інвагінації цитоплазматичної мембрани частина протопласта відділяється від материнської клітини. Цей протопласт містить частину ядерного матеріалу — один геном. Утворення клітинної стінки між обома протопластами (як за звичайного поділу клітини) не відбувається. Замість цього протопласт чохлика. Екзоспорій міститься тільки у не багатьох бактерій (наприклад у Bacillus cereus). майбутньої спори оточується, мовби обростає, плазматичною мембраною материнської клітини. В результаті навкруги протопласта розміщуються дві плазматичні мембрани і кожна з них бере участь у синтезі стінки спори. Мембрана протопласта спори синтезує назовні від себе стінку зародкової клітини, а мембрана, яка походить від материнської клітини, синтезує всередину кору спори (кортекс). Кортекс складається з багатошарового пептидогліканового остова, який відрізняється від каркаса стінок вегетативної клітини ступенем зшивання. Зовнішню оболонку спори утворює материнська клітина. Ця оболонка складається з поліпептидів. Материнська клітина утворює також ще один додатковий поліпептидний шар — екзоспорій, який оточує спору у вигляді особливого чохлика.Єкзоспорій містеться у не багатьох бактерій.

Рис. 2.11. Спороутворення і будова зрілої спори:
А, Б — процес відділення протопласта спори; В, Г,Д — утворення передспори; Е — зріла спора; 1 — цитоплазма; 2 — плазматична мембрана; 3 — клітинна стінка зародкової клітини; 4 — кора спори; 5,6 — відповідно внутрішня і зовнішня оболонки спори; 7 — екзоспоріум
Індукція спороутворення. Спори не є обов'язковою стадією життєвого циклу бактерій. За сприятливих умов бактерії можуть необмежений час розмножуватися поділом, як вегетативні клітини. Утворення спор починається лише тоді, коли не вистачає поживних речовин або коли в надлишку накопичуються продукти обміну. Інакше кажучи, воно відбувається лише тоді, коли для цього створюються певні умови. Висихання не стимулює спороутворення. Якщо помістити вегетативні клітини в дистильовану воду, можна спостерігати "ендотрофну споруляцію", тобто утворення спор за рахунок внутрішньоклітинних запасних речовин. У таких випадках утворення спор, очевидно, викликане браком екзогенного субстрату. Індукція спороутворення відбувається впродовж декількох годин. Якщо, наприклад, до суспензії вегетативних клітин Bacillus cereus у перші 5 год після внесення їх у дистильовану воду добавити глюкозу, то утворення спор припиниться: добавлення субстрату пригнічує споруляцію. Але якщо глюкозу добавити пізніше, ніж через 6 год, пригнічення спороутворення не відбувається. Індукція спороутворення продовжується, і вже через 10-13 год після внесення у воду близько 90 % клітин утворюють спори. Отже, споруляція регулюється зовнішніми факторами.Кількість клітин, що утворюють спори, у багатьох випадках збільшується після добавлення в середовище солей марганцю.
Здатність утворювати ендоспори поступово втрачається за багаторазових пересівів вегетативних клітин. Оскільки суспензія спороутворювальних бактерій містить і спори, і вегета-
тивні клітини, перед кожним пересіванням культуру, як правило, піддають короткочасному кип'ятінню. Це сприяє зберіганню чи підвищеній здатності клітин утворювати спори.
Властивості зрілих спор. Спори вивільнюються при автолізі материнських клітин. Зрілі спори не виявляють ніякої метаболічної активності. Вони надзвичайно стійкі до дії високих температур, різного роду опромінень, хімічних агентів. Терморезистентність зумовлена низьким вмістом води у спорах — 15 % (стільки само, скільки в сухому казеїні або у вовні) і приблизно пропорційна вмісту в них дипіколінової кислоти.
Проростання спор. У відповідних сприятливих умовах більшість спор проростають. Відповідне попереднє оброблення, певні умови зберігання та прогрівання можуть підвищити "схожість" спор — збільшити процент проростання. У випадку Bacillus subtills оптимальними умовами для стимуляції проростання спор вважають семиденний період спокою і п'ятихвилинне прогрівання у воді при 60 °С. Інші спори можуть бути активовані короткочасним кип'ятінням (10 хв, 100 °С). Теплове оброблення спор повинно проводитися безпосередньо перед висіванням, оскільки процес активації є зворотним.
Проростанню спор передує поглинання ними води та набухання. Для проростання активованих спор у ряді випадків необхідною є наявність глюкози, амінокислот, нуклеозидів та інших речовин. У ході проростання відбуваються глибокі фізіологічні зміни: дихання та ферментативна активність швидко підвищуються, починається виділення амінокислот, дипіколінової кислоти та пептидів. У процесі проростання втрати сухої речовини досягають 25-30 %. Під час проростання спори втрачають термостійкість. Ростова трубка, яка виходить із спори, оточена дуже тонкою, не до кінця сформованою клітинною стінкою, так що в протопласт може проникати навіть ДНК. Ростова трубка може утворюватись як у полярному, так і латеральному положенні {рис.2.12). В одному випадку оболонка спори при цьому розривається, в іншому — ростова трубка її проколює.
Тривалість життя спор. Бактерії у вигляді спор можуть тривалий час перебувати у стані анабіозу. Так, у зразках ґрунту, що зберігалися 50-100 років, були виявлені спори бацил. За даними такого роду експериментів, у сухому ґрунті за 50 років зберігання до 90 % спор втрачають життєздатність. Але при цьому 1 т сухого ґрунту навіть через 1000 років міститиме життєздатні спори.
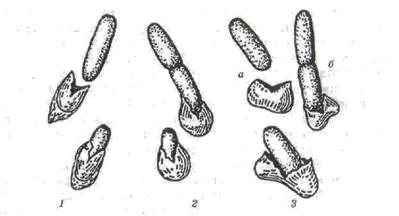
Рис. 2.12. Проростання спори:
1,2 — полярне; 3 — латеральне; а — Bacillus cereus; б — Bacillus subtilis
У сухому стані багато бактерій (якщо не всі з них) упродовж багатьох років зберігають життєздатність. Для консервації бактерій у колекціях культур вегетативні клітини, як правило, піддають ліофільному висушуванню (висушуванню з замороженого стану) і зберігають при кімнатній температурі або низьких температурах у вакуумі. Бактерії, які не витримують ліофілі-зації, зберігаються упродовж багатьох років у вигляді суспензії при температурі рідкого азоту.
3.3. Інші форми спокою (цисти, екзоспори, міксоспори)
Крім ендоспор, у деяких бактерій спостерігається утворення інших форм спокою — екзоспор і цист. Екзоспори утворюють метаноокиснювальні бактерії роду Methylosinus і фототрофна пурпурова бактерія Rhodomicrobium vannielii. Екзоспори утворюються брунькуванням материнської клітини. Екзоспори значно менш термостійкі, ніж ендоспори, але стійкіші до висушування, ультрафіолетового опромінення, ніж вегетативні клітини. Екзоспори не містять дипіколінової кислоти, вони не стійкі до дії лізоциму, погано фарбуються різними барвниками.
Деякі бактерії утворюють кулеподібні товстостінні клітини — цисти,. Цисти містять цитоплазму з нуклеоїдом (і гранулами полі-в-гідроксимасляної кислоти у Azotobacter vinelandli), оточену цитоплазматичною мембраною та двома оболонками. Виникають у старих культурах за відсутності поживних ресурсів, при цьому в цисту перетворюється вся клітина. На відміну від вегетативних клітин, цисти містять багато ліпідів. Цисти стійкіші до висушування, механічних пошкоджень, дії лізоциму, ніж вегетативні клітини, але малорезистентні до температури. У сприятливих умовах (за наявності джерел вуглецю) цисти проростають. Цисти утворюють бактерії родів Azotobacter, Bdellovlbrio, Methylococcus, спірохети. У метанотрофних бактерій роду Methylococcus цисти утворюються в несприятливих умовах, наприклад, за браку кисню. У багатьох випадках цистоутворення у цих бактерій супроводжується зміною пігментації клітин від жовтого до темно-коричневого кольору. Культура, яка не утворює цист, залишається кремовато-білою. Але у Methylococcus thermophilus цистоутворення не завжди супроводжується посиленням пігментації.
У міксобактерій утворення цист, які називаються міксоспорами, є закономірною стадією їх життєвого циклу. Вегетативні клітини міксобактерій після закінчення стадії активного бінарного поділу за умов дефіциту джерел живлення збираються разом у масу і формують різної форми плодові тіла, які складаються із слизу та міксоспор. При цьому тільки частина клітин перетворюється на міксоспори, решта гине, лізується і є будівельним матеріалом для утворення плодових тіл.
Міксоспори мало відрізняються від вегетативних клітин, але є такі, що сильно різняться за формою, стійкістю до висушування, УФ-опромінення, температури, наявністю додаткових оболонок. Такі міксоспори називаються мікроцистами.
У деяких прокаріот спори є одночасно і формами спокою, і репродуктивними системами. У актиноміцетів родів Thermoactinomyces та Actinobifida спостерігається ендогенне утворення спор, але у більшості родів актиноміцетів спори формуються екзогенно — артроспори, конідієспори та ін. Артроспори формуються виникненням поперечних перетинок у гіфах з відділенням сегментів так, що утворюється ланцюжок спор. Конідієспори утворюються на поверхні особливих гіфів конідієносців, або конідієфорів. Спорангіоспори — спори, що формуються в спорангіях. Оскільки спорангіоспори утворюються всередині сі рангіів, їх називають ендогенними. Спорангії утворюються на гіфах — спорангіофорах.
Форми спокою деяких ціанобактерій називають акінетами. Вони більші ніж вегетативні клітини, мають подовжену або сферичну форму товсту оболонку. Утворення акінет відбувається в період затримки росту культури. Клітина при цьому збільшується в розмірах, у цитоплазмі накопичуються гранули запасних речовин (глікоген, поліфосфати, ціанофіцинові гранули), а також має місце утворення карбоксисом. Одночасно потовщується пептидоглікановий шар, ущільнюється слизовий чохол. В цілому, оболонки акінет мають більший вміст ліпідів і полісахаридів, а цитоплазма —менше води, ніж вегетативна клітина. У процесі формування акінет збільшується вміст ДНК, кількість рибосом, зменшується кількість хлорофілу і фікобілінових пігментів.
Форми спокою прокаріот характеризуються низьким рівнем метаболічної активності, підвищеною стійкістю до дії несприятливих факторів, здатністю тривалий час перебувати у життєздатному стані..
|
|
|
|
Дата добавления: 2014-01-06; Просмотров: 7638; Нарушение авторских прав?; Мы поможем в написании вашей работы!