КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Генетика поведения Dictyostelium discoideum
|
|
|
|
Dictyostelium discoideum (диктиостелиум) — это клеточный слизевик, относящийся к типу Mycetozoa. Описанный в 1935 году диктиостелиум вскоре стал одним из интереснейших модельных организмов в клеточной биологии, генетике и биологии развития.
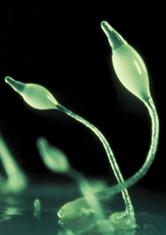
Рис.4.4. Dictyostelium discoideum - модельный организм в клеточной биологии, генетике и биологии развития.
Большую часть жизни диктиостелиум проводит в виде одиночных почвенных амеб, однако при определенных условиях эти амебы образуют подвижные агрегаты, а затем многоклеточные плодовые тела сложного строения. Происходящие при этом процессы межклеточной сигнализации, клеточной дифференцировки и морфогенеза позволяют использовать его в качестве модельного объекта.
В природе Dictyostelium discoideum обитает в почве и подстилке (влажном листовом опаде). Одиночные амёбы Dictyostelium discoideum питаются в основном бактериями - их привлекает выделяемая бактериями фолиевая кислота.
Жизненный цикл диктиостелиума начинается с того, что его споры высвобождаются из зрелых плодовых тел и разносятся ветром. При достаточно высокой влажности и температуре из спор выходят миксамёбы — одноклеточная стадия развития диктиостелиума. При достаточной влажности и наличии пищи они питаются и делятся с помощью митоза.
При исчерпании пищи начинается агрегация миксамёб. На этой стадии в клетках миксамёб синтезируются специфические гликопротеиды и аденилатциклаза. Гликопротеиды обеспечивают межклеточную адгезию, а аденилатциклаза синтезирует цАМФ.

Рис.4.5. Образование цАМФ
цАМФ, секретируемый в среду, служит, как и у бактерий, сигналом «клеточного голода». У диктиостелиума цАМФ также является хемоаттрактантом для голодных миксамёб. Несколько случайно оказавшихся радом и «склеившихся» первыми миксамёб служат центром, к которому привлекаются и ползут со всех сторон голодные миксамёбы. Соединяясь с помощью молекул клеточной адгезии, они образуют агрегат из нескольких десятков тысяч клеток.
Изначально плоский агрегат совершает сложные движения, приподнимается над субстратом, а затем ложится на бок и превращается в мигрирующего «слизня» — подвижный псевдоплазмодий длиной 2-4 мм.
Псевдоплазмодий мигрирует всё время одним концом вперед, и в его составе начинается дифференцировка клеток. Часть клеток на переднем конце образуют полисахаридную оболочку. Псевдоплазмодий мигрирует сквозь неё, и какая-т ее часть остается на субстрате в виде слизистого следа.
Псевдоплазмодий движется по направлению к свету, более высокой температуре и большей сухости воздуха.
цАМФ и вещество, известное как фактор индукции дифференцировки (DIF), стимулируют дальнейшую дифференцировку и формирование нескольких типов клеток.
На переднем конце псевдоплазмодия оказываются клетки — предшественники стебелька плодового тела, а на заднем — клетки-предшественники спор. После окончания миграции в подходящих условиях клетки псевдоплазмодия совершают сложные перемещения и формируют соответствующие части плодового тела.
«Сходные с передними» (аnterior-like) клетки, открытые сравнительно недавно, распределены по задней половине тела псевдоплазмодия. Эти клетки формируют подставку (самую нижнюю часть плодового тела).
После остановки псевдоплазмодия за счет перемещения клеток формируются «стадия сомбреро», а затем начинается кульминационная фаза формирования плодового тела.
В ходе этой фазы передние и задние клетки псевдоплазмодия меняются местами. Передние клетки «сомбреро» формируют целлюлозные клеточные стенки и собираются в полый трубчатый стебелек, по наружной поверхности которого клетки — предшественницы спор мигрируют вверх, а остальные клетки — предшественницы стебелька — вниз.
Полностью сформированное плодовое тело высотой 1-2 мм образуется через 8-10 часов после начала этой фазы. После созревания спор на его вершине споры рассеиваются, и цикл начинается снова.
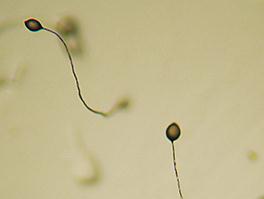
Рис.4.6. Плодовое тело Dictyostelium discoideum
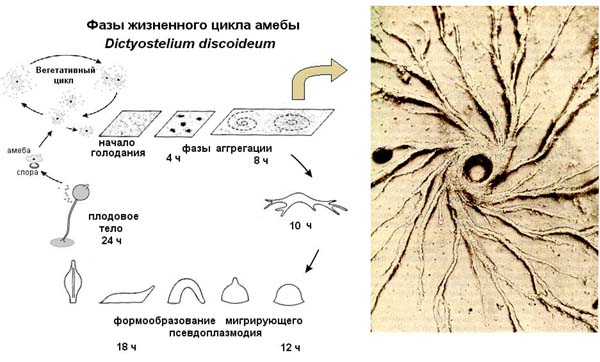
Рис.4.7. Жизненный цикл Dictyostelium discoideum
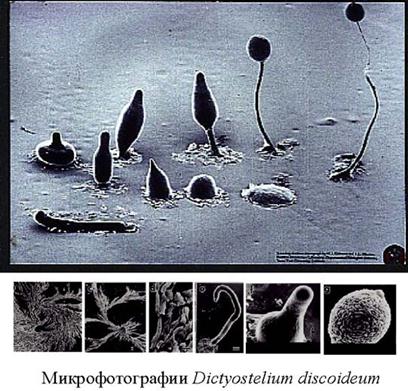
Рис.4.8. Микрофотографии жизненного цикла Dictyostelium discoideum
Кроме описанной выше бесполой части цикла, в жизненном цикле диктиостелиума может происходить и половое размножение. Переход к половому размножению может запускаться подсыханием подстилки, где живут миксамёбы. Сливаясь, две миксамёбы разных типов спаривания образуют зиготу — «гигантскую клетку». Зигота начинает заглатывать окружающих миксамёб. Проглотив несколько сотен миксамёб, зигота выделяет толстую целлюлозную оболочку, формируя так называемую макроцисту.
Макроциста делится сначала с помощью мейоза, а затем с помощью митоза, формируя множество гаплоидных миксамёб. Выходя из-под оболочки макроцисты, они начинают питаться и размножаться бесполым путём. Таким образом, в жизненном цикле Dictyostelium discoideum единственная диплоидная стадия — это зигота.
Преимущества диктиостелиума как модельного объекта — относительно простое строение, небольшое число типов клеток, а также короткий жизненный цикл и простота выращивания в лабораторных условиях.
Хемотаксис у Dictyostelium discoideum изучается на примере движения миксамёб по направлению к источнику секреции цАМФ. В секреции цАМФ и скорости передвижения миксамёб наблюдается цикличность с определенным периодом. Интересно, что использование цАМФ в качестве хемоаттрактанта не описано ни у одного другого организма. В прокариотических и животных клетках цАМФ служит внутриклеточным сигналом, а Dictyostelium discoideum – единственный организм, у которого цАМФ действует еще и как внеклеточная сигнальная молекула
При формировании плазмодия отдельные клетки слизевика агрегируют в результате хемотаксиса. Одна из реакций на голодание у миксамеб состоит в том, что они начинают вырабатывать и выделять цАМФ, который служит хемотаксическим сигналом, привлекающим других миксамеб. Агрегация инициируется случайным образом: любые клетки, начинающие первыми секретировать цАМФ, привлекают другие клетки и таким образом становятся центрами агрегации. цАМФ, вырабатываемый такими «клетками-инициаторами», секретируется отдельными «импульсами» и связывается специфическими рецепторами на поверхности соседних голодающих амеб, направляя тем самым их движение в сторону источника цАМФ.
Такой хемотаксический ответ можно продемонстрировать, нанеся из микропипетки ничтожное количество цАМФ на любой участок поверхности клетки голодающей миксамебы. Ответом будет немедленное образование псевдоподии, растущей в сторону пипетки. Псевдоподия прикрепляется к поверхности, на которой находится клетка, и тянет клетку в том же направлении. Как только образуется центр агрегации, зона его влияния быстро расширяется, так как агрегирующие клетки не только отвечают на сигнал цАМФ, но и передают его от клетки к клетке.
Каждый импульс цАМФ побуждает соседние клетки не только к движению к источнику импульса, но и к испусканию собственного импульса в виде цАМФ. Этот новый, высвобождающийся с небольшой задержкой импульс в свою очередь ориентирует находящиеся рядом клетки, вызывая у них тоже выброс цАМФ, и т.д.
Таким образом возникают регулярные чередующиеся волны цАМФ, распространяющиеся из каждого центра агрегации, заставляя более удаленных миксамеб двигаться внутрь концентрическими или спиральными волнами. Преимущество такой системы передачи состоит в том, что по мере распространения из центра сигнал постоянно возобновляется, не ослабляясь на большом расстоянии.
Помимо активации сигнальной системы цАМФ голодание миксамеб Dictyostelium вызывает экспрессию сотен новых генов, и некоторые из них кодируют молекулы межклеточной адгезии, участвующие в агрегации клеток.
Амёбы Dictyostelium в последние годы стали излюбленным объектом биологов, изучающих эволюцию кооперации и социального поведения. Ученых интересуют скопления одноклеточных индивидуумов, которые создают идеальные условия для развития социального паразитизма и нахлебничества. Для того чтобы выжить, социальным организмам вроде диктиостелиума необходимо каким-то образом защищаться от нахлебников.
Исследования на диктиостелиуме, показали, что вероятность развития устойчивости в результате случайных мутаций у этого организма довольно высока. Ученые работали с двумя штаммами диктиостелиума — «честными» амёбами дикого типа (условное обозначение штамма — AX4) и одним из нескольких известных штаммов амёб-«обманщиков» (chtC). Если смешать амёб из этих штаммов в равной пропорции и начать морить их голодом, они образуют химерные (смешанные) плодовые тела. При этом «обманщики» занимают лучшие места в плодовом теле и превращаются в споры, предоставляя «честным» амёбам в одиночку строить ножку плодового тела. В результате среди образовавшихся спор резко преобладают споры обманщиков.
Ученые искусственно повысили темп мутирования у «честных» амёб AX4 при помощи генетических конструкций (плазмид), встраивающихся в различные участки генома диктиостелиума. В состав плазмиды входил ген устойчивости к антибиотику бластицидину S. Встраиваясь в разные места генома, эта плазмида влияла на работу близлежащих генов. Затем из множества получившихся амёб-мутантов взяли тысячу особей с разными мутациями и каждой из них дали возможность размножиться.
После этого стали вести отбор на устойчивость к нахлебникам, причем в качестве отбирающего агента использовались сами нахлебники. Амёб из тысячи мутантных штаммов смешивали в равной пропорции и объединяли с амёбами-обманщиками, причем последних было в четыре раза больше, чем честных амёб-мутантов. Смешанную популяцию морили голодом, заставляя образовывать плодовые тела. Затем собирали образовавшиеся споры и выводили из них амёб. Естественно, среди них преобладали обманщики chtC, но экспериментаторы убивали их всех бластицидином S (ведь все амёбы-мутанты имели ген, защищающий их от этого антибиотика). В результате получалась смесь амёб-мутантов, но из тысячи исходных штаммов в ней теперь преобладали те, кто смог лучше других противостоять обманщикам. Этих амёб снова смешивали с обманщиками в пропорции 1: 4 и снова заставляли образовывать плодовые тела.
После шести таких циклов в популяции амёб-мутантов остались представители только одного из тысячи исходных штаммов. Авторы исследовали геном этих амёб и выяснили, что плазмида у них встроилась в ген DDB_G0271758, кодирующий белок с неизвестной функцией.
Выживший мутантный штамм назвали rccA (resister of cheater chtC A).
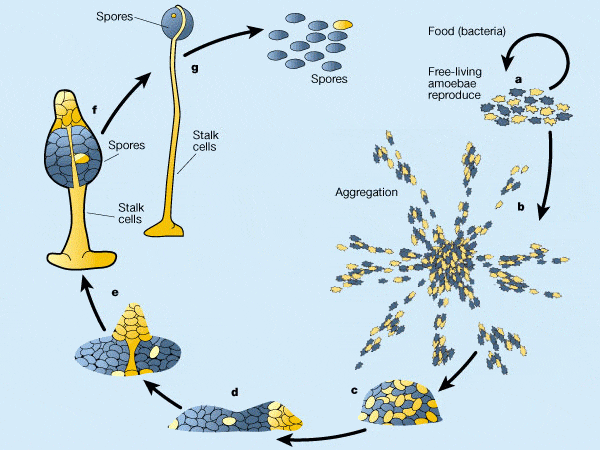
Рис.4.9. Социальный паразитизм у Dictyostelium
а — при избытке пищи амёбы живут поодиночке, растут и размножаются бесполым путем (делением); b–c — при недостатке пищи амёбы собираются в большие скопления. d — в результате образуются многоклеточные агрегаты длиной в несколько миллиметров. e–g —многоклеточный агрегат превращается в «плодовое тело» на ножке; при этом около 20% клеток жертвуют собой, образуя ножку (желтые клетки - «честные»), а 80% превращаются в споры и получают шанс продолжить свой род (синие клетки - «обманщики»).
Авторы убедились, что амёбы rccA действительно защищены от нахлебничества со стороны обманщиков-chtC. Если смешать тех и других в равной пропорции, то споры в химерных плодовых телах образуются тоже в равной пропорции — следовательно, жульнические приемы амёб chtC, в чём бы они ни заключались, бессильны против амёб rccA. Однако другой штамм амёб-обманщиков, LAS1, успешно паразитировал на амёбах rccA. Следовательно, мутация в гене DDB_G0271758 защитила амёб не от любых обманщиков, а только от вполне определенных.
Исследователи также проверили, не стал ли устойчивый штамм rccA сам «обманщиком» по отношению к исходному штамму AX4. Теоретически, один из способов одолеть обманщика в эволюционной «гонке вооружений» — это самому стать еще более искусным обманщиком. Подобное соревнование между обманщиками в итоге может привести всю систему, основанную на кооперации, к полному краху. Однако в данном случае этого не произошло: штамм rccA остался вполне «честным» по отношению к AX4.
Эксперименты со смешанными культурами, состоящими из равного количества амёб AX4 («диких»), chtC («обманщиков») и rccA («защищенных»), показали, что амёбы rccA защищают от обмана не только себя, но и диких амёб AX4 (хотя и в несколько меньшей степени). Присутствие амёб rccA каким-то образом мешает обманщикам chtC вытеснять амёб AX4 из выгодных позиций в плодовых телах. Ясно, что взаимопомощь честных штаммов открывает дополнительные возможности для борьбы с обманщиками.
Исследование показало, что вероятность появления мутаций, обеспечивающих защиту от нахлебников, у диктиостелиума весьма высока. Присутствие нахлебников способствует распространению защитных мутаций. Это должно приводить к эволюционной «гонке вооружений» между обманщиками и честными амёбами: первые совершенствуют средства обмана, вторые — средства защиты. Для того чтобы защититься от обманщиков, амёбы не обязаны сами становиться обманщиками. Это способствует сохранению кооперации.
Генетические механизмы обмана и защиты от него пока остаются неизвестными, но скорее всего они связаны с системами межклеточной коммуникации и взаимного узнавания.
Глава 5. Генетика поведения беспозвоночных животных.
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 2111; Нарушение авторских прав?; Мы поможем в написании вашей работы!