КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Генетические основы нейрогуморальной регуляции поведения насекомых
|
|
|
|
Эндокринные железы и нейросекреция активно влияют на процессы роста и развития насекомых. Деятельность всех желез внутренней секреции, с самого их возникновения регулировалась центральной нервной системой по нервным проводникам, продуктами нейросекреции или посредством гормонов других эндокринных желез, выведение которых в кровь происходило в результате нервных импульсов. Поэтому гормональная и нервная регуляция образуют единый регуляторный механизм.
У всех многоклеточных животных, начиная с плоских червей, регуляцию и интеграцию всех функций организма осуществляет центральная нервная система. Именно она обеспечивает ответные реакции целостного организма на все воздействия внешней или внутренней среды, вызывающие раздражения рецепторов. Воздействия центральной нервной системы на эффекторы могут осуществляться двумя путями: путем передачи импульсов возбуждения по эфферентным нервам (нервно-проводниковый путь) и путем введения в кровь или лимфу гормонов и других физиологически активных веществ (гуморальный путь).
Гуморальный путь реализуется с помощью желез внутренней секреции, которые образованы скоплениями железистого эпителия, пронизанного большим количеством кровеносных и лимфатических сосудов, а также нервных окончаний. Выделяемые ими гормоны оказывают регулирующее влияние на определенные ткани или органы. Для осуществления действия гормона необходимо выполнение следующих условий: осуществление синтеза гормона, его активация (созревание), доставка к месту «работы» и наличие в органах или тканях, на которые оказывается влияние клеток-мишеней со специфическими рецепторами данного гормона.
Деятельность желез внутренней секреции подвержена ритмическим колебаниям, как суточным, так и сезонным. Например, колебания активности половых желез в связи с циклами размножения. Периодичность в нарастании уровня тех или иных гормонов способствует формированию в ЦНС очагов возбуждения - доминант, «запускающих» инстинктивное поведение.
Гормоны могут действовать на все отделы нервной системы, некоторые безусловные рефлексы могут осуществляться только при достаточном содержании определенных гормонов.
Влияние нервной системы на функционирование желез внутренней секреции может осуществляться как опосредованно, через изменение концентрации определенных веществ, влияющих на эти железы, так и непосредственно, путем нервной регуляции. Например, у самок тараканов особые эндокринные железы, так называемые прилежащие тела, вырабатывают гонадотропный гормон, стимулирующий рост ооцитов в яичниках. Однако нервные импульсы, идущие из мозга, тормозят функцию прилежащих тел и секрецию этого гормона. Погружение сперматофора в совокупительную сумку самки во время спаривания рефлекторно прекращает тормозящее действие нервных импульсов на прилежащие тела, что ведет к выведению в гемолимфу гормона, вызывающего рост ооцитов и поступление в них желтка.
Генетические основы нейрогуморальной регуляции полового поведения насекомых.
Для большинства насекомых половое поведение связано, прежде всего, с поиском полового партнера. При наличии соответствующего гормонального фона двигательная активность животных на этапе такого поиска резко усиливается. Но локомоторные реакции, сопровождающие поиск полового партнера, приближение к нему, удаление от него и другие, являются обычными двигательными реакциями.
Выбор брачного партнера — одна из самых ответственных задач в жизни любого насекомого, способного к такому выбору. Системы коммуникации, позволяющие животным выбирать оптимальных партнеров, бывают весьма сложными. Они могут включать много разных типов сигналов:
· звуковые (песни)
· зрительные (брачная окраска)
· поведенческие (специальные позы, танцы)
· хеморецепторные (различные запахи, феромоны, а также специфические молекулы, несущие персональную информацию)
При этом смысл каждого сигнала может зависеть от контекста, то есть от других сигналов, что крайне затрудняет для исследователей их расшифровку.
В химической коммуникации насекомых важную роль играют углеводородные феромоны. Например, Drosophila melanogaster производит десятки разных углеводородов, различающихся длиной углеродной цепи, ее разветвленностью, числом и положением двойных связей. Все они, по-видимому, участвуют в химическом общении мух, однако о значении конкретных молекул пока известно очень немногое.
Было выяснено, что одни углеводороды производятся преимущественно самками, другие — самцами. «Мужской» углеводород 7-T (Z-7-трикозен) привлекает самок, но отпугивает самцов. «Женские» углеводороды 7,11-HD (7Z,11Z-гептакозадиен) и 7,11-ND (7Z,11Z-нонакозадиен), по-видимому, привлекают самцов. Генетически модифицированные самцы, выделяющие эти женские феромоны, воспринимаются нормальными самцами как самки и подвергаются сексуальным домогательствам.
Было установлено, что самцы во время спаривания передают самкам порцию феромона cVA (cis -vaccenyl acetate), который отпугивает других самцов и повышает вероятность того, что данная самка сохранит верность своему первому партнеру.
Основную помощь в поисках партнера многим насекомым оказывает обоняние. Воспринимают запахи насекомые с помощью плакоидных, триходиных, базиконических и целоконических сенсилл. (Рис. 5.15., 5.16., 5.17).
Сенсиллы (от лат. sensilis — чувствующий) — простейшие кожные органы чувств у беспозвоночных, выполняющие роль органов осязания, вкуса, обоняния. Все сенсиллы возникли из одного первоначального типа, путем специализации для восприятия определённого раздражения.
Сенсилла представляет собой видоизмененный участок покровов тела (волосок, ямка, пластинка и т. п.), к которому подходят отростки одного или нескольких чувствительных нейронов. Наружная часть образована клетками покровов и представлена волоском (осязательные сенсиллы) либо тонкой кутикулярной пластинкой, часто в форме купола или конуса, погруженной в специальную ямку (обонятельные и вкусовые сенсиллы).
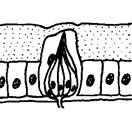
Рис. 5.15. Плакоидные обонятельные сенсиллы
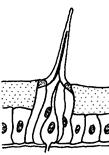
Рис. 5.16. Триходиные обонятельные сенсиллы
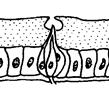
Рис. 5.17. Целоконические обонятельные сенсиллы
У некоторых видов насекомых для восприятия запахов служат особые опоясывающие нити, чувствительные конусы, кутикулярные выросты и еще более сложные образования. Все эти дистантные хеморецепторы располагаются почти исключительно на антеннах.
Так, например, рецепторная часть обонятельной системы имаго дрозофилы включает две пары головных придатков (Рис. 5.18). Около 1200 ольфакторно-чувствительных (ольфакторный от лат. olfactorius-обонятельный) чувствительных нейронов локализованы на каждом из третьих антеннальных сегментов, причём они ассоциированы в кластеры по 2–4 нейрона, морфологически обособленные как ольфакторные сенсиллы (волоски).
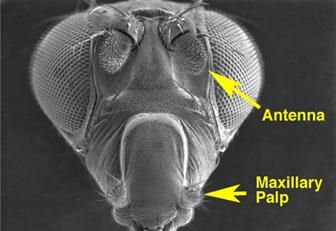
Рис.5.18. Органы обоняния дрозофилы
Активация ольфакторно чувствительных нейронов происходит при взаимодействии феромона или другого летучего вещества с одним из ольфакторных рецепторных белков (OR), который экспрессируется на поверхности дендрита.
От каждого ольфакторно-чувствительного нейрона отходит раздвоенный аксон, ветви которого оканчиваются в соответствующих зонах двух антеннальных долей головного мозга – первичном центре обработки обонятельной информации. Затем проекционные нейроны проводят нервный импульс к латеральным рожкам и грибовидным телам, представляющим собой высший нервный центр, где и происходит окончательный анализ ольфакторной информации и её трансформация в поведенческий ответ.
Чувствительность насекомых ко многим запахам того же порядка, что и у человека, иногда - несколько выше, иногда — ниже. Однако чувствительность насекомых к половым аттрактантам может быть крайне высокой. Например, самец тутового шелкопряда способен различать половой аттрактант самки своего вида – бомбикол – в концентрации 100 молекул на 1 см3.. Самец грушевой сатурнии способен различать феромоны самки на расстоянии 5-10 км.
Такая сверхчувствительность насекомых к специфическим запахам предполагает очень строгую дифференциацию ольфакторных стимулов. Аксоны соответствующих сенсорных нейронов проводят нервные импульсы в обонятельные доли дейтероцеребрума (это второй, или средний, отдел головного мозга, иннервирующий антенны) непосредственно, без синаптических переключений.
Следует отметить, что половые феромоны Drosophila не являются летучими веществами; они экскретируются на поверхность кутикулы и регистрируются особями противоположного пола уже в процессе выполнения ритуала ухаживания при помощи контактных хеморецепторов, расположенных на ротовых органах и лапках.
Эксперименты с генетически модифицированными мухами Drosophila melanogaster, лишенными способности производить углеводородные феромоны, позволили частично расшифровать химический язык насекомых, при помощи которого мухи выбирают брачных партнеров.
Одна из главных трудностей в изучении этого химического языка насекомых состоит в том, что экспериментаторам крайне трудно манипулировать составом углеводородов, находящихся на поверхности кутикулы.
Чтобы создать мух, не способных производить углеводородные феромоны, ученые вставили в геном дрозофилы особую генетическую конструкцию, которая должна была уничтожить эноциты — специализированные клетки, располагающиеся под кутикулой брюшка у насекомых. Считается, что именно эноциты отвечают за синтез углеводородных феромонов. Поскольку эноциты являются модельным объектом, используемым для изучения генетической регуляции индивидуального развития, то известны промоторы (регуляторные участки генов), активизирующиеся только в эноцитах и нигде более.
Чтобы получить мух без эноцитов, ученые вставили в геном дрозофилы ген Wrinkled, активизация которого заставляет клетку совершить апоптоз, объединенный с «эноцитовым» промотором. В результате в ходе развития насекомого клетки, превратившиеся в эноциты, вскоре погибают.
Сначала все генетически модифицированные насекомые погибали на стадии личинки. Вероятно, личинки дрозофил не могут жить без эноцитов. Генетическую конструкцию усложнили так, чтобы ген апоптоза включался только у взрослых мух, а у личинок оставался неактивным. После этого ученым удалось получить живых взрослых дрозофил, лишенных эноцитов (oe–).
И самцы, и самки oe– оказались почти полностью лишены углеводородных феромонов. Тем самым была подтверждена гипотеза о том, что именно эноциты отвечают за их производство. Исключение составил мужской феромон cVA, который производится не эноцитами, а клетками половой системы.
Мух oe– использовали для изучения роли отдельных углеводородов. Выяснилось, что отсутствие углеводородных феромонов не приводит к видимым изменениям брачного поведения (ухаживания). Самцы oe– совершенно нормально ухаживали за самками дикого типа, лишь немного уступая в активности «диким» самцам. Правда, особым успехом у самок они не пользовались: в среднем у самца oe– уходило вчетверо больше времени на соблазнение самки, чем у дикого самца. Таким образом, отсутствие феромонов у самца почти не влияет на его собственное поведение, но снижает его привлекательность для самок.
Став малоинтересными для самок, «дезодорированные» самцы oe– при этом стали весьма привлекательными для самцов (как диких, так и других oe–). Судя по всему, самцы oe– воспринимаются другими самцами как самки. Взаимные ухаживания самцов oe– приводят к причудливым трансформациям полового поведения. Например, два самца могут долго кружиться, уткнувшись друг в друга головами.
Самец oe– перестает быть объектом гомосексуальных домогательств, если обработать его мужским феромоном 7-T. Это подтверждает предполагавшуюся ранее роль 7-Т в предотвращении гомосексуальных взаимодействий у самцов дрозофил.
Дезодорированные самки oe– с точки зрения диких самцов стали совершенно неотразимыми («сексуальными гиперстимулами»). При наличии выбора самцы предпочитали спариваться с такими самками, а не с обычными, выделяющими разнообразные углеводороды. Это значит, что женские углеводородные феромоны служат не для привлечения самцов, а наоборот — для их сдерживания. Возможно, это позволяет самке в более спокойной обстановке разобраться, подходит ли ей этот самец.
Вероятно, феромоны могут содержать информацию о половой и видовой принадлежности, а их влияние на сексуальную привлекательность может быть разным в зависимости от контекста. Чтобы проверить эти предположения, ученые обрабатывали «дезодорированных» мух oe– женским феромоном 7,11-HD (который до сих пор считался «женским афродизиаком», то есть средством привлечения самцов) и мужским феромоном cVA, при помощи которого самцы отпугивают от своей партнерши других ухажеров. Ранее было показано, что феромон cVA действительно снижает привлекательность самок, однако действие его не абсолютно: многие самки в природе всё-таки откладывают яйца, оплодотворенные несколькими разными самцами.
Оказалось, что сам по себе женский феромон 7,11-HD не влияет на привлекательность самок oe–. Однако он существенно уменьшает отпугивающий эффект феромона cVA. Самки oe–, обработанные феромоном cVA, не вызывали интереса у самцов. Однако если самку oe– обработать одновременно обоими феромонами — 7,11-HD и cVA — то она становится такой же привлекательной для самцов, как и дикие самки.
По-видимому, сексуальная привлекательность самки определяется сочетанием сигналов-репеллентов, при помощи которых самец пытается отвратить от нее других самцов, и сигналов, выделяемых самкой, которые подавляют действие репеллентов. В этом проявляется конфликт интересов самцов и самок. Самцы заинтересованы в верности своих партнерш, а самкам выгодно иметь более широкий выбор генов для своего потомства, поэтому они предпочитают спариться с несколькими самцами.
Исследователям удалось показать роль углеводородных феромонов в формировании межвидовой репродуктивной изоляции. Для этого скрещивали D. melanogaster (диких и oe–) с мухами из трех близких видов: D. simulans, D. yakuba и D. erecta. Из этих трех видов у двух первых вообще нет специфических женских углеводородных феромонов, а у третьего вида они есть, но не такие, как у D. melanogaster.
Самцы всех трех видов либо вовсе не хотели ухаживать за дикими самками D. melanogaster, либо делали это очень неохотно. Однако они охотно ухаживали за самками oe–. Следовательно, углеводородные феромоны, выделяемые самками D. melanogaster, участвуют в поддержании репродуктивной изоляции между видами: они отпугивают «чужих» самцов.

Рис.5.18.а. Роль углеводородных феромонов в формировании межвидовой репродуктивной изоляции.
Верхний ряд: лишенные эноцитов и кутикулярных углеводородов самцы и самки являются «сексуальными гиперстимулами» для диких самцов D. melanogaster, а также для самцов трех близких видов.
Средний ряд: обработка одиночными феромонами снижает привлекательность мух oe–.
Нижний ряд: добавление второго феромона подавляет эффект первого, и в результате привлекательность мух снова возрастает.
Функцию «отпугивания» может выполнять женский феромон 7,11-HD. Если самку oe– обработать этим феромоном, она перестает привлекать самцов чужих видов. Более того, если тем же феромоном обработать самку D. simulans, D. yakuba или D. erecta, она становится непривлекательной даже для своих родных самцов. Зато самка любого из четырех видов, обработанная этим феромоном, становится желанным партнером для самцов D. melanogaster. Самцы D. melanogaster проявляют интерес к самкам D. erecta даже без предварительной обработки феромоном 7,11-HD. Возможно, это объясняется тем, что самки D. erecta, как и самки D. melanogaster, производят другой специфический женский феромон — 7,11-ND.
Таким образом, феромон 7,11-HD информирует самцов не только о половой, но и о видовой принадлежности самок D. melanogaster.
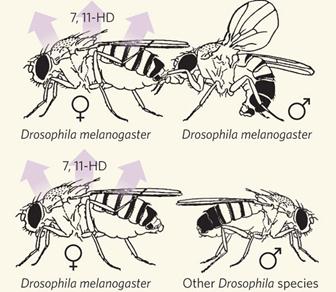
Рис.5.19. Феромон 7,11-HD информирует самцов о видовой принадлежности самок D. Melanogaster. Самки D. melanogaster выделяют углеводородный феромон — вещество 7,11-HD (7,11-гептакозадиен), которое отпугивает самцов других видов, но не «своих».
У разных видов дрозофил одни и те же углеводородные феромоны могут выполнять разные функции. Например, феромон T-7 у D. melanogaster вырабатывается преимущественно самцами, привлекает самок и отпугивает других самцов. У D. simulans этот феромон вырабатывается в большей степени самками и не отталкивает, а привлекает самцов. Оказалось, что если намазать самку oe– феромоном T-7, это делает её лишь немного более привлекательной для самцов D. simulans. Если такую же самку намазать феромоном 7,11-HD, то это делает ее неинтересной для самцов всех видов, кроме «своего». Однако феромон T-7 обладает способностью резко уменьшать отталкивающее действие феромона 7,11-HD на самцов D. simulans. Иными словами, эффективность феромона T-7 зависит от того, была ли самка oe– предварительно обработана феромоном 7,11-HD: если да, то феромон Т-7 резко повысит ее привлекательность для самцов D. simulans, если нет — эффект будет гораздо слабее.
Очевидно, что возможность сложных взаимодействий между феромонными сигналами характерна не только для D. melanogaster. Вероятно, эта возможность заложена в общей архитектуре системы коммуникации при помощи углеводородных феромонов и соответствующих рецепторов.
Логика феромонной коммуникации у дрозофил основана на том, что все мухи исходно являются сексуально привлекательными друг для друга (по крайней мере для самцов), и на эту общую исходную привлекательность феромоны накладывают «дополнительную» информацию о половой и видовой принадлежности. При этом смысл отдельных сигналов может зависеть от контекста, включающего множество других сигналов: химических, зрительных, звуковых и прочих.
Как происходит передача феромонального сигнала?
Предполагают наличие двух основных молекулярных компонентов –феромон-связывающего белка (PBP – pheromone binding protein) и продукта одного из генов Or (Olfactory receptor) семейства, - при помощи которых происходит передача феромонального сигнала. В качестве феромон-связывающего белка, опосредующего действие агрегационного феромона дрозофилы как цис-вакценил ацетат (cis-vaccenyl acetate), известен LASH – белок.
Мухи с функционально неактивным геном lash демонстрируют пониженную агрегационную активность. Более того, по электрофизиологическим записям lash - мутантов установлено, что нейроны трихоидных сенсилл, в норме экспрессирующих LASH, у этих мутантов не стимулируются под воздействием цис-вакценил ацетата. Искусственное же введение LASH в трихоидную сенсиллу таких насекомых приводит к полному восстановлению цис-вакценил ацетат опосредованных электрических ответов. Эти данные свидетельствуют о необходимости данного белка для активации ольфакторно чувствительных нейронов, экспрессирующих специфический рецептор к комплексу цис-вакценил ацетат/LASH.
Агрегационное и пищевое поведение представляют собой очень похожие поведенческие реакции – движение по направлению к желаемому источнику запаха, но биологическое значение каждой из этих реакций уникально. Для самцов и виргинных (неоплодотворенных) самок агрегационное поведение – это способ повышения шансов спариться. Для оплодотворённых самок агрегационные феромоны – это сигнал мест яйцекладки, используемых другими самками. Благодаря такому явлению как агрегационное поведение имаго для личинок, выходящих из яиц, создаются оптимальные условия роста. Т.е. создаётся оптимальная плотность популяции, что важно для качества пищи, низкого уровня смертности и нормальной скорости развития.
Распознавание агрегационного феромона должно вовлекать те же самые центральные мозговые структуры, которые активируются при распознавании пищевых аттрактантов. Вероятно последующие поведенческие реакции (яйцекладка, спаривание) должны инициироваться другими феромонами, поскольку мухи достигли источника агрегационного феромона, то есть, обнаружение источника агрегационных феромонов является завершающим актом соответствующего поискового поведения.
Большую помощь в поисках полового партнера многим насекомым оказывает зрение. Своеобразная окраска ряда бабочек, ночное свечение жуков-светляков и т. п. — все это в большинстве случаев признаки и сигналы, служащие для опознания и привлечения особей своего вида.
Насекомые, обладающие хорошо развитыми органами слуха, нередко производят для привлечения полового партнера те или иные звуки. Это - сверчки, кузнечики, цикады и многие др.
В определенные периоды полового цикла все основные органы чувств насекомых служат одной цели: найти особь противоположного пола своего вида. Все сигналы для встречи особей противоположного пола обладают, как правило, высокой видовой специфичностью, а система обнаружения этих сигналов — поразительной помехоустойчивостью.
Интересен нейро–гормональный контроль ритуала ухаживания. Ухаживание самцов представляет собой комплекс врождённых поведенческих актов, вовлекающих многие сенсорные модальности: зрительную, контактную хеморецепцию и звуковую рецепцию. Последовательность стадий репертуара ухаживания, а, следовательно, и порядок вовлечения тех или иных сенсорных систем не является случайной.
В общих чертах схема нейрологического контроля репертуара ухаживания у D. melanogaster представлена на Рис.5.20.
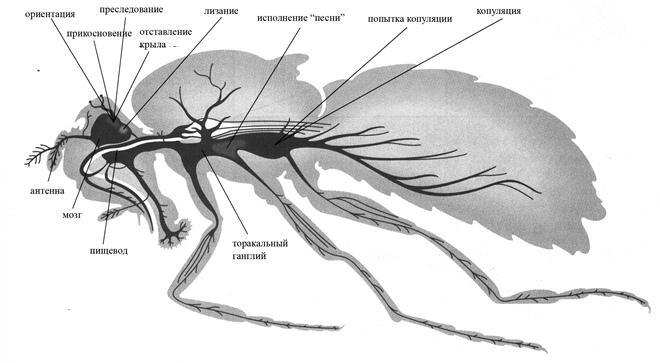
Рис. 5.19. Схема нейрологического контроля отдельных стадий репертуара ухаживания дрозофилы.
На первой стадии ухаживания – стадии ориентации – ключевой сенсорной модальностью является зрительная. Основными органами зрения у насекомых на стадии имаго служат сложные глаза, дополнительными – дорсальные (теменные) глазки. Структурной и функциональной единицей фасеточного глаза насекомых является омматидий (от греч. ōmma, род. падеж ōmmatos — глаз). Омматидии различаются у разных отрядов насекомых.
Поля зрения у мух перекрываются лишь на уровне зрительных центров (в первом оптическом ганглии – в ламине). Такие глаза получили название нейросуперпозиционных. Вероятно, таким путём у мух достигается повышение абсолютной чувствительности глаза без увеличения размера фасеток и снижения остроты зрения.
Фоторецепторы насекомых передают сигналы в зрительные центры без помощи нервных импульсов, электротонически. Большинство исследователей полагают, что сигналом в фоторецепторах служат рецепторные потенциалы, имеющие достаточную амплитуду и способные электротонически градуально активировать зрительные интернейроны.
Преобразование зрительной информации начинается уже на уровне фоторецепторов, каждый из которых отбирает для передачи в высшие центры лишь определённый признак стимула. Кроме того, у некоторых насекомых между фоторецепторами одного омматидия могут наблюдаться электрические связи, обеспечивающие суммацию потенциалов определённых рецепторов и способствующие тем самым лучшему выделению полезного сигнала из общего набора сигналов.
Дальнейшая и более глубокая обработка зрительной информации осуществляется на уровне оптических ганглиев.
Сигналы от фоторецепторов, оканчивающихся в I оптическом ганглии, поступают на вход униполярных нейронов I порядка. Полагают, что аксоны фоторецепторов в месте синаптических переключений выделяют какой-то тормозной медиатор, поскольку в униполярных нейронах в ответ на освещение глаза возникают гиперполяризационные потенциалы. Как и в фоторецепторах, потенциалы действия в интернейронах I порядка не генерируются, и, таким образом, эти нейроны электротонически передают сигналы к другим интернейронам зрительных центров.
На униполярных интернейронах конвергируют сигналы от нескольких фоторецепторов, в результате чего здесь происходит суммация этих сигналов, и светочувствительность интернейронов I порядка оказывается выше по сравнению с рецепторами.
Важную роль в первичной обработке информации на уровне I оптического ганглия играет и латеральное торможение, которое наиболее выражено между соседними нейромматидиями. «Геометрия» тормозных латеральных связей специфична у разных видов насекомых, что позволяет им наиболее эффективно выделять в изображении значимые именно для них признаки.
Сведения о локальных признаках объекта, даваемые нейромматидиями, подлежат дальнейшей обработке и анализу. За счёт горизонтальных (тангенциальных) интернейронов эти сведения суммируются, причём имеются интернейроны, образующие синаптические связи с самым различным числом нейромматидиев и даже со всеми неромматидиями ганглия.
Функциональные задачи, решаемые такими суммирующими интернейронами II и более высокого порядка, могут быть самыми разнообразными. Установлено, что в зрительной системе насекомых имеются нейроны-детекторы, способные выделять из всего объёма зрительной информации лишь определённые признаки стимулов или сочетаний таких признаков. Нейроны-детекторы найдены не только в оптических ганглиях, но и в средней части переднего мозга. Например, обнаружены нейроны-детекторы движения, высокочастотные детекторы движения, нейроны-детекторы формы. Помимо этих узкоспециализированных зрительных интернейронов у насекомых обнаружены интернейроны «общего» действия, реагирующие на изменения освещённости.
У насекомых обнаружены полимодальные интернейроны, реагирующие как на зрительную, так и на какую-либо иную стимуляцию, а также интернейроны, не относящиеся непосредственно к зрительной системе, но способные модулировать чувствительность и активность зрительных интернейронов. С помощью такого рода нейронов зрительный анализатор насекомого включается в целостную систему регуляции поведения животного.
Переход к следующей стадии репертуара ухаживания, т.е. генерация двигательного акта в соответствии с полученной сенсорной информацией, по-видимому, осуществляется при участии комплекса центрального тела. Именно туда приходят аксоны из самых различных частей мозга, и эта структура обеспечивает непосредственные связи с подглоточным ганглием и ганглиями туловищного мозга.
Репертуар ухаживания D. melanogaster включает две феромон-опосредованные стадии: стадия прикосновения и стадия лизания, поскольку в данном случае феромоны не являются летучими веществами. Следовательно, для их распознавания необходим тесный физический контакт или, по крайней мере, максимальное приближение источника феромонов на брюшке самки и рецепторного нейрона, локализованного на ноге самца, если речь идёт о стадии прикосновения, или на лабеллярных пальпах, в случае стадии лизания. Феромоны D. melanogaster синтезируются в эноцитах и затем транспортируются в эпидермальные клетки кутикулы, откуда секретируются на её поверхность.
Специфический для самцов феромон во время спаривания переносится на поверхность брюшка самки, и, по-видимому, функционирует как один из ингибиторов ухаживания по отношению к ней со стороны других самцов.
Гормональная регуляция синтеза феромонов у насекомых осуществляется преимущественно посредством трёх основных соединений: ювенильного гормона, 20-гидроксиэкдизона и нейрогормона пептидной природы – PBAN (pheromone biosynthesis activating neuropeptide). У D. melanogaster гормональная регуляция выработки феромонов изучена не очень подробно, вероятно в ней принимают участие ювенильный гормон, 20-гидроксиэкдизон и некий пока не установленный пока мозговой фактор.
Восприятие половых аттрактантов самцами Drosophila происходит при помощи контактных хеморецепторов (вкусовых рецепторов). Первичным вкусовым органом является лабеллум (губа) – пара пальп в дистальной части пробосциса (хоботка). Пальпа содержит 31 вкусовую сенсиллу – щетинки, – каждая из которых состоит из одного механо – и 2-4 хемосенсорных нейронов, окружённых вспомогательными клетками.
Вкусовые сенсиллы подразделяют на различные морфологические классы: S (small) – малые; I (intermediate) – средние; L (large) – крупные.
Дополнительные хемочувствительные клетки, обнаруженные на медиальной поверхности каждой пальпы и на внутренней поверхности глотки, по-видимому, в большей степени имеют отношение к питанию и контактируют непосредственно с пищей при её поглощении. У Drosophila вкусовые щетинки есть также на лапках и по краю крыла. Именно они имеют непосредственное отношение к процессу ухаживания, количество вкусовых щетинок на передних лапках у самцов значительно выше, чем у самок. У самцов – 50, а у самок - 37. Кроме того, нейроны, ассоциированные с этими щетинками экспрессируют специфический для самцов gustatory receptor (Gr) ген, продукт которого функционирует как рецептор феромонов.
Вкусовые рецепторные нейроны (подобные ольфакторно чувствительным нейронам) – это сенсорные нейроны I типа, объединённые во вкусовые сенсиллы и экспрессирующие вкусовые рецепторы – GRs (gustatory receptors). Эти рецепторы специфично взаимодействуют с растворимыми веществами, например, сахарами, аминокислотами и другими.
Все вкусовые рецепторные нейроны лабиальных пальп и некоторые, локализованные на лапках, проецируют аксоны в подглоточный ганглий мозга – первичный центр обработки вкусовой информации, но аксоны некоторых вкусовых рецепторных нейронов, локализованных на лапках, заканчиваются в грудном ганглии.
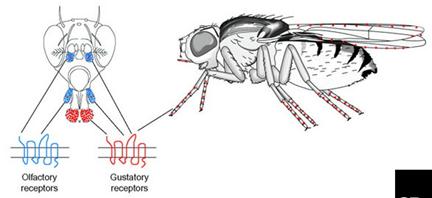
Рис. 5.20. Преимущественная локализация обонятельных и вкусовых сенсилл у дрозофилы
Ещё одна стадия репертуара ухаживания, о регуляции которой следует упомянуть – стадия вибрации. У Drosophila отсутствуют специализированные звукопроизводящие органы. Звуки же, издаваемые самцами дрозофилы на этой стадии, являются побочным следствием работы мышц крыла. Управление системой звукопроизведения со стороны нервной системы состоит в запуске этих мышц и поддержании их работы в определённом ритме.
Работой мышц крыла насекомых непосредственно управляют ганглии грудного отдела нервной цепочки. Однако рецептивные поля, участвующие в активации нейронов этих мышц, находятся на значительном удалении от ганглиев, на определённых участках тела насекомого. На большом числе видов насекомых (стрекозах, тараканах, саранче, осах, пчёлах, бабочках, мухах, жуках) было показано, что именно конечности являются той инстанцией, которая опосредует запуск и прекращение летательных движений.
По-видимому, система инициации полёта у дрозофилы адаптирована к выполнению ухаживания.
Сегментарные ганглии насекомых содержат все необходимые механизмы, обеспечивающие даже очень сложные формы поведения, но эта сегментарная координационная система не способна осуществлять адекватную реакцию насекомого на изменения во внешней среде без участия высших центров.
Наиболее важными центрами регуляции локомоторной активности являются грибовидные тела и центральное тело, связанные между собой антагонистическими взаимоотношениями, а также подглоточный ганглий.
Надглоточный и подглоточный ганглии, не определяя большей частью картины той или иной формы мышечной активности, регулируют её уровень при помощи тормозных или возбуждающих неспецифических влияний. Иными словами, высшие локомоторные центры модулируют типы движений, определяемые сегментарными ганглиями, обеспечивая тем самым поведение насекомого как единого организма.
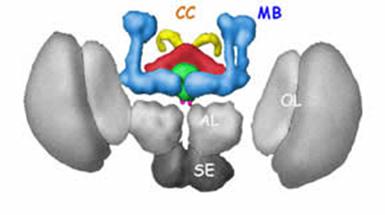
Рис. 5.21. Схематическое трёхмерное изображение головного мозга дрозофилы:
MB (mushroom bodies) – грибовидные тела; red, fan shaped bodies; green, ellipsoid bodies; pink, noduli; yellow, protocerebral bridge.
CC (central complex) – центральный комплекс, включающий все четыре вышеупомянутых элемента;
AL (antennal lobes) – антеннальные доли; SE (subesophageal ganglia) – подглоточный ганглий;
OL (optic lobes) – оптический доли.
Восприятие звуков, в частности, у двукрылых осуществляется с помощью джонстоновых органов, которые представляют собой наиболее сложно устроенные хордотональные органы и локализуются во втором членике антенн.
По своему строению, эти органы сходны у разных видов насекомых. В состав каждой сенсиллы джонстонова органа входят 3 сенсорных нейрона и 3 обслуживающих клетки; дендриты нейронов несут на вершинах по одной видоизменённой ресничке, покрытой снаружи кутикулярной оболочкой. Вершина этой оболочки прикрепляется к сочленовной мембране, соединяющей второй и третий членики антенны.
Фонорецепторы джонстонова органа реагируют не на абсолютную величину звукового давления, как у позвоночных животных, а на его изменения – градиент давления. Импульсы возбуждения передаются нейронами джонстонова органа непоредственно в головной мозг, где и находятся несколько высших центров анализа слуховой информации.
По-видимому, слуховые центры функционально связаны и с грибовидными телами, поскольку переход к следующей стадии ухаживания – стадии лизания – сопровождается снижением подвижности самок, а именно грибовидные тела являются основным источником тормозных влияний на локомоторную активность насекомых.
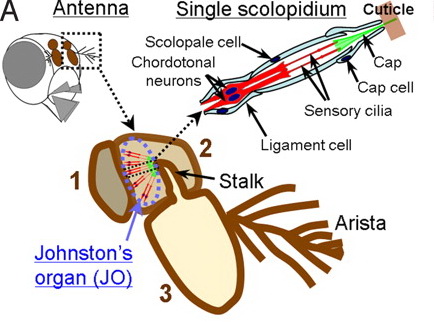
Рис.5.22. Схематическое изображение расположения и строения джонстонова органа дрозофилы.
В управлении поведенческими реакциями у насекомых, равно как и у других высокоорганизованных животных, существуют различные уровни, т.е. иерархия командных систем управления. Подобное разделение функций внутри центральной нервной системы биологически обосновано, поскольку освобождает головные ганглии (главным образом надглоточный) для выполнения наиболее сложных функций, связанных с условнорефлекторной деятельностью, памятью и др. Для насекомых это особенно важно, так как число нейронных элементов, включённых в интегративную деятельность, у них относительно невелико.
Несмотря на некоторую фрагментарность знаний по нейрофизиологии внутренних органов насекомых, очевидно, что управление ими со стороны нервной системы является достаточно сложным, что это управление осуществляется в тесном взаимодействии с гуморальными факторами. В отличие от управления локомоторной активностью наибольшее значение здесь приобретает вегетативная нервная система.
Последовательность событий при управлении половым поведением насекомых в самых общих чертах такова. Все изменения во внешней среде животное улавливает с помощью соответствующих рецепторов. Информация об этих изменениях передается в мозг, где она анализируется, и с помощью эндокринных органов (прилежащих и кардиальных тел) создает необходимый (для данного этапа) гормональный фон, обеспечивая готовность насекомого к осуществлению той или иной стадии полового цикла. Однако для реализации определенных двигательных реакций, сопровождающих половое поведение, необходимы дополнительные сведения об окружающей обстановке, например, о наличии полового партнера и т. п. Эти сведения насекомое также получает с помощью органов чувств. Сигнал передается в мозг, который запускает соответствующие реакции, придавая им, если нужно, ту или иную гормональную окраску. Таким образом, непосредственная регуляция полового поведения насекомых обеспечивается сложным комплексом нейрогуморальных воздействий.
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 982; Нарушение авторских прав?; Мы поможем в написании вашей работы!