КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Кинетические модели Эйгеновского типа
|
|
|
|
ДОБИОЛОГИЧЕСКАЯ МОЛЕКУЛЯРНАЯ ЭВОЛЮЦИЯ
• Способность генных сетей к самовоспроизведению является фундаментальным свойством, обретенным в ходе добиологической молекулярной эволюции. Однако очевидно, что основные понятия биологической теории эволюции: особь, наследование, наследственная изменчивость, генотип, фенотип, популяция и т.д., в общем случае неприменимы к добиологической эволюции. Поэтому элементарным объектом добиологической эволюции следует считать фракцию макромолекул (полинуклеотидов, полипептидов). От других фракций она должна отличаться последовательностью мономеров и молекулярной функцией.
• Соответственно, в добиологической эволюции элементарной эволюционирующей единицей является устойчивый коллектив (ансамбль) макромолекул, т.е. совокупность фракций макромолекул, которые связаны между собой процессами синтеза (матричного и нематричного), ферментативного катализа, стереохимического взаимодействия, деградации, модификации и т.д.
• Сеть функциональных взаимодействий ансамбля и набор концентраций его молекулярных компонент образует динамическую структуру ансамбля. Эволюцией ансамбля собственно и будет являться изменение его динамической структуры.
• Подобного рода устойчивые ансамбли макромолекул с одной стороны имеют свойства особей (стабильная внутренняя структура, самовоспроизведение) и с другой стороны популяций (способность эволюции путем мутаций и отбора). Устойчивость системы самовоспроизведения определяется ее циклическим характером.
В 1973 г. М. Эйген применил химико-кинетический подход к описанию динамики коллективов информационных макромолекул. Эти модели позволяют описать возможные варианты организации коллективов макромолекул и оценить с эволюционных позиций их свойства, хотя это, вероятно, и не относится к реальному эволюционному процессу
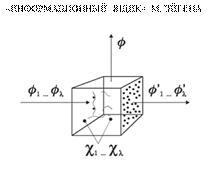
• Будем считать, что процессы матричного синтеза макромолекул далеки от равновесия и протекают только в комплексах матриц (полинуклеотидных) и ферментов, осуществляющих этот синтез. Обозначим xi — полную концентрацию макромолекул i-го типа, a zi — концентрацию комплекса (комплексов), производящего макромолекулы i-ro типа.
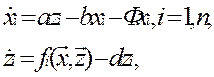 |
• где ai — параметр синтеза, bi — параметр распада макромолекул i-ro типа, а Ф — единый поток разбавления всех макромолекул, fi — функция формирования комплексов i -го типа, di — константа их дезинтеграции.
• Примем так называемое «селекционное ограничение» — дополнительное требование (в некотором смысле – граничное условие), связанное с регулированием потока Ф. Будем использовать только ограничение, обеспечивающее так называемое постоянство внутренней организации коллектива макромолекул, а именно:
• а) концентрации мономеров нуклеиновых кислот и белков, а также других субстратов в резервуаре избыточны, т. е. параметры а i, b i, d i и функции f i и Ф не зависят от них;
• 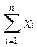 б) суммарная плотность макромолекул С постоянна, т.е.
б) суммарная плотность макромолекул С постоянна, т.е.

что достигается регулированием потока разбавления Ф. Это ограничение обусловлено тем, что реальные системы, где может идти синтез макромолекул — клетки, коацерваты — растут или дробятся не переуплотняясь, а это эквивалентно регулированию стока;
• в) механизм разбавления одинаков для всех фракций, что описывается членом -Ф xi.
• Кроме того, Эйген предполагает, что комплексы zi квазиравновесны, т. е. равновесие по ним устанавливается значительно быстрее, чем изменяются x i и поток разбавления. Тогда, полагая dz/dt = 0, получаем систему алгебраических нелинейных уравнений:
dizi 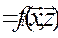 , i=1,n
, i=1,n
Введенная динамическая система для коллектива макромолекул оказывается полностью эквивалентна уравнениям популяционной генетики. Введем относительные концентрации
ui = xi/C
и величины
wi = aizi/xi – bi
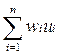 которые являются аналогами «селективных ценностей», или приспособленностей отдельных фракций макромолекул.
которые являются аналогами «селективных ценностей», или приспособленностей отдельных фракций макромолекул.
Усредняя по коллективу, получаем w =
• 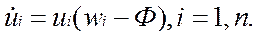 Это — аналог средней приспособленности коллектива. Тогда система уравнений динамики приобретает хорошо известный из генетики популяций вид
Это — аналог средней приспособленности коллектива. Тогда система уравнений динамики приобретает хорошо известный из генетики популяций вид
Суммируя по i, получаем, что
w = Ф
т. е. поток разбавления имеет смысл средней приспособленности коллектива макромолекул. Из популяционной генетики известно, что эта величина определяет темп размножения популяции.
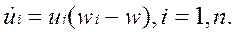 Поэтому естественно требовать, чтобы w = Ф > 0 и окончательно получаем
Поэтому естественно требовать, чтобы w = Ф > 0 и окончательно получаем
• — аналог уравнений популяционной генетики с отбором. Очевидно, что фракции, для которых wi < w, уменьшают относительные концентрации, а если wi > w, то фракции макромолекул растут. Достижение равновесия возможно при достижении внутренне сбалансированного состояния коллектива макромолекул, иногда оно наступает после вытеснения некоторых фракций, имеющих относительную приспособленность меньше средней.
• Однако, в отличие от популяционно-генетических задач, приспособленности здесь не введены как параметры, а получены из кинетических констант и динамических переменных после задания молекулярных процессов в коллективе. Величины wi в общем случае не константы, а нелинейные функции частот ui. Поэтому аналог фундаментальной теоремы естественного отбора Фишера (dw/dt≥0) в общем случае не выполняется. Если коллективы внутренне сбалансированы и внешне обособлены от других фракций макромолекул, то их можно считать автономными единицами отбора следующего уровня, пересчитав приспособленности на эти коллективы.
• Возможны и более сложные режимы функционирования описываемой системы.
• Основная проблема, возникающая в связи с анализом систем самовоспроизведения макромолекул, — это проблема их структурной и динамической устойчивости и внутренней сбалансированности.
• Один из способов решения этой проблемы - циклический характер системы самовоспроизведения.
• Эйген предложил вариант циклического коллектива, состоящего из полинуклеотидов и полипептидов (с разделением функций между ними) и назвал эту конструкцию каталитическим гиперциклом с трансляцией. Матрица Ii воспроизводится с помощью белка Ei -1 и эта матрица кодирует белок Ei, который необходим для воспроизведения матрицы Ii+1. Таким образом, гиперцикл по сути дела является обобщением каталитического цикла белков с отдельной матрицей для каждого звена. При этом подразумевается, что уже существует система генетического кодирования белков и трансляции (Ii → Ei). На рисунке она не показана.
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 432; Нарушение авторских прав?; Мы поможем в написании вашей работы!