КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Світлова фаза фотосинтезу
|
|
|
|
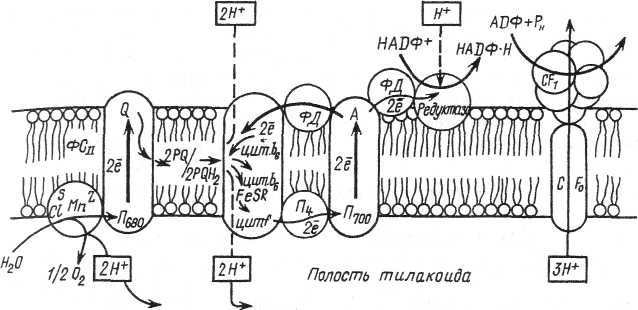
Суть світлової фази фотосинтезу складається в поглинанні променистої енергії і її трансформації в асиміляційну силу (АТФ і НАДФ·Н2), необхідну для відновлення вуглецю в темнових реакціях. Складність процесів перетворення світлової енергії в хімічну вимагає їх строгої мембранної організації. Світлова фаза фотосинтезу відбувається в гранах хлоропласта.
До складу фотохімічно активних мембран хлоропластів входять п'ять узгоджено функціонуючих багатокомпонентних білкових комплексів: світлозбираючий комплекс, фотосистеми I і II, цитохромний комплекс (цитохроми b6 і f), що бере участь у транспорті електронів, і АТФ-азний комплекс, що забезпечує синтез АТФ.
Організація й функціонування пігментних систем. Пігменти хлоропластів об'єднані у функціональні комплекси – пігментні системи, у яких реакційний центр – хлорофіл а, що здійснює фотосенсибілізацію, пов'язаний з процесами переносу енергії антеною, що складається зі світлозбираючих пігментів. Сучасна схема фотосинтезу вищих рослин включає дві фотохімічні реакції, здійснювані за участю двох різних фотосистем (ФСІ та ФСІI).
До складу ФСІ як реакційний центр входить димер хлорофілу а з максимумом поглинання світла 700 нм (Р700), а також хлорофіли а 675-695, що грають роль антенного компонента. Первинним акцептором електронів у цій системі є мономерна форма хлорофілу а 695, вторинними акцепторами – залізосірчані білки (-FeS). Комплекс ФСІ під дією світла відновлює залізовмісний білок – фередоксин (Фд) і окислює мідьвмісний білок – пластоціанін (Пц).
ФСІІ включає реакційний центр, що містить хлорофіл а (Р680) і антенні пігменти – хлорофіли а 670-683. Первинним акцептором електронів є феофітин (Фф), що передає електрони на пластохінон. До складу ФСІІ входить також білковий комплекс S-системи, що окислює воду, і переносник електронів Z. Цей комплекс функціонує за участю марганцю, хлору й магнію. ФСІІ відновлює пластохінон (PQ) і окислює воду з виділенням О2 і протонів.
Сполучною ланкою між ФСІІ й ФСІ служать фонд пластохінонів, білковий цитохромний комплекс (b6 і f), і пластоціанін.
У хлоропластах рослин на кожен реакційний центр доводиться приблизно 300 молекул пігментів, які входять до складу антенних або світлозбираючих комплексів. Антенний білковий комплекс ФСІ містить 110 молекул хлорофілів а 680-695 на один Р700, з них 60 молекул – компоненти антенного комплексу, який можна розглядати як світлозбираючий білковий комплекс (ССК) ФСІ. Антенний комплекс ФСІ також містить ß-каротин.
Антенний білковий комплекс ФСІІ містить 40 молекул хлорофілів а з максимумом поглинання 670-683 нм на один Р680 і ß-каротин.
Хромопротеїни антенних комплексів не мають фотохімічної активності. Їхня роль складається в поглинанні й передачі енергії квантів на невелику кількість молекул реакційних центрів Р700 і Р680 кожна з яких пов'язана з ланцюгом транспорту електронів і здійснює фотохімічну реакцію. Організація електронно-транспортних ланцюгів (ЕТЛ) при всіх молекулах хлорофілу нераціональна, тому що навіть на прямому сонячному світлі на молекулу пігменту кванти світла попадають не частіше одного разу за 0,1 с.
Передача енергії у світлозбираючих комплексах і фотосистемах відбувається шляхом індуктивного резонансу. Умовами для резонансного переносу енергії служать зближене розташування й перекриття частот осцилюючих електронних систем у двох взаємодіючих молекул. Причому міграція енергії здійснюється від короткохвильових пігментів до більш довгохвильових. Квант світла мігрує у фотосистемі, поки не потрапить у стокову молекулу (пастку) – пігмент реакційного центра, що є самим довгохвильовим і тому нездатним збуджувати сусідні молекули. Енергія збудженого пігменту реакційного центра витрачається на фотохімічну роботу. Квант світла звичайно передається на стокову молекулу через кілька сотень передач, кожна передача здійснюється за 10-12 с, 100 передач – відповідно за 10-10 с. Експериментально встановлено, що 90 % енергії збудження хлорофілу а може бути передане на Р700 менш ніж за 10-9 с.
Нециклічне і циклічне фотосинтетичне фосфоритування.
Фотосинтетичне фосфорилювання, тобто утворення АТФ у хлоропластах у ході реакцій, що активуються світлом, може здійснюватися циклічним і нециклічним шляхами.
Циклічне фотофосфорилювання є більш простим й еволюційно більш древнім. При циклічному фотофосфорилюванні функціонує тільки ФСІ й її єдиним продуктом є АТФ.
Циклічне фотофосфорилювання було відкрито в 1954 р. Арноном, Алленом і Френкелем на ізольованих хлоропластах шпинату.
|
При поглинанні кванта світла один з електронів пігменту реакційного центра (Р700) переходить на більш високий енергетичний рівень. У цьому стані він захоплюється білком, що містить залізо й сірку (Fe-S-центр), а потім передається на залізовмісний білок фередоксин. Подальший шлях електрона – поетапний, його транспорт назад до Р700 через ряд проміжних переносників, серед яких є флавопротеїди й цитохроми. Під час транспорту електрона його енергія вивільняється й використовується на приєднання фосфору неорганічного (Фн) до АДФ із утворенням АТФ. Кількість молекул АТФ, що утворюються при переносі одного електрона, дотепер не встановлено.
Отже, при циклічному фотофосфорилюванні енергія світла витрачається на перенос електрона на високовідновлене з'єднання, тобто проти градієнта ОВ-потенціалу, а потім електрон «скочується з гірки», утвореної переносниками з рівнем відновленості який знижується, і, збіднений енергією, повертається на ФСІ. Механізми сполучення (АТФ-азний комплекс) забезпечують на цьому відрізку шляху запасання енергії у вигляді АТФ. У найпростіших автотрофів (бактерій) – це єдиний шлях фотофосфорилювання.
У вищих рослин у процесі еволюції з'явився більш складний шлях, що здійснюється при участі двох фотосистем і забезпечує відновлення НАДФ за рахунок фотоокислення води. Причому відновлення НАДФ здійснює ФСІ, а фотоокислення води – ФСІІ. Ці дві системи функціонують одночасно й взаємозалежно. Р. Хіллом і Ф. Бендаллом (1960) розроблена схема послідовності реакцій, що одержала назву схеми нециклічного транспорту електронів, або Z-схеми.
При збудженні Р700 в реакційному центрі ФСІ електрон захоплюється мономерною формою хлорофілу а і потім послідовно передається через залізосірчані білки, фередоксин, флавопротеїди на відновлення НАДФ. Р700 не одержавши електрона назад, як у випадку циклічного фосфорилювання, набуває позитивний заряд, що компенсується електроном ФСІІ.
У ФСІІ Р680 збуджений квантом світла, передає електрон феофітину. Від феофітина електрон, втрачаючи енергію, послідовно передається на пластохінони, залізосірчаний білок, цитохром f, пластоціанін і, нарешті, на Р700 ФСІ. Енергія, що вивільняється при транспорті електрона від збудженої ФСІІ на ФСІ, використовується для синтезу АТФ із АДФ і Фн, тобто тут має місце фотофосфорилювання.
Р680 залишившись без електрона, набуває здатність одержувати електрон від води. Незважаючи на активне дослідження, детально механізм процесу фотоокислення води не встановлений. Показано участь білкового комплексу й переносника електронів Z, для функціонування яких необхідні Mn, Cl і Са.
Таким чином, при нециклічному шляху відбувається лінійний або відкритий, тобто не замкнутий по циклу, транспорт електронів. Донором електронів є вода, кінцевим акцептором – НАДФ. Причому відбувається одночасно двоелектронний транспорт. Передача електронів здійснюється при участі двох фотосистем, тому для переносу кожного електрона витрачаються два кванти світла. На ділянці між ФСІІ і ФСІ транспорт електрона йде по зниженню градієнта окисно-відновного потенціалу з вивільненням енергії й запасанням її в АТФ.
Поряд з нециклічним у мембранах хлоропластів вищих рослин функціонує циклічний транспорт електронів. При цьому фередоксин виконує роль регулятора потоку електронів. При зростанні потреби в АТФ частина електронів від фередоксину через систему цитохромів повертається до Р700 з утворенням АТФ. Відновлення НАДФ у цьому випадку не відбувається, і фотоокислення води при участі ФСІІ не потрібно.
Білкові комплекси, що беруть участь у фотохімічних реакціях фотосинтезу, в мембранах хлоропластів розподілені нерівномірно. Найбільші розходження спостерігаються між тими мембранами, які щільно зв’язані одна з одною у гранах, і тими ділянками мембран, які контактують зі стромою, — закруглені ділянки тилакоїдів гран і мембрани тилакоїдів строми. Світлозбираючий комплекс і ФСІІ перебувають в основному в щільнозістикованих мембранах гран. ФСІ зі своїм світлозбираючим комплексом й АТФ-азний комплекс перебувають на незістикованих ділянках мембран. Взаємодія між ними здійснюється за допомогою легкорухомого в ліпідній фазі пластохінона, а також завдяки переміщенню водорозчинного пластоціаніна уздовж внутрішніх поверхонь ламел і водорозчинного фередоксина уздовж зовнішніх поверхонь.
До латеральних переміщень здатні й пігмент-білкові комплекси. Наприклад, відновлення в надлишкових кількостях пластохінона ФСІІ сприяє переміщенню ССК у стромальну область мембрани й зростанню міграції поглиненої енергії світла із ФСІ. Підвищена фотохімічна активність ФСІ підсилює окислювання пластохінона, що приводить до зворотного переміщення ССК. Таким чином, система взаємної регуляції активності ФСІ і ФСІІ включає латеральний транспорт білкових комплексів у мембранах хлоропластів.
Синтез АТФ. Передача електрону по ЕТЛ проходить за градієнтом окис¬но-відновного потенціалу і супроводжується вивільненням енергії, яка використовується для синтеза АТФ. Синтез АТФ відбувається за рахунок електрохімічного градиента протонів, що накопичуються у полості тилакоїду грани.
Існує декілька теорій, що пояснюють механізм фосфорилювання АДФ, поєднаний з транспортом електронів. Найбільшим визнанням користується хеміосмотична теорія англій¬ського біохіміка П. Мітчела (1961). Згідно цієї теорії, пластохінон, приєднав два електрони від П680, приєднує ще два протони зі строми хлоропласта і переносить їх крізь мембрану до внутритилакоїдного простору. Протони накопичуються всередині тилакоїда і в результаті відбувається фотоокислення води.
Завдяки нерівномірному розподілу протонів по обидві сторони мембрани утворюється різниця хімічних потенціалів протонів, і виникає електрохімічний мембранний потенціал іонів водню (∆μН+). ∆μН+ включає дві складові: концен¬траційну (∆рН), що виникає внаслідок нерівномірного розподілу протонів по обидві сторони мембрани, і електричну (∆ψ), що обумовлена виникненням протилежного за¬ряду на поверхні мембран, тобто утворенням мембранного потенціалу. Енергія ∆рН і ∆ψ використовується для зворотного транспор¬ту протонів з внутритилакоїдного простору в строму хлоропласта по особливим каналам. З зворотним транспортом протонів спряжено фосфорилювання АДФ.
Спряження зворотного транспорту протонів і фосфорилювання АДФ забезпечує Н+-АТФ-синтетаза, що розташована в тилакоїдних мембранах і складається з двох частин: водорозчинної каталітичної частини (F1), зверненої до строми хлоропласта, і мем¬бранної частини (F0), що пронизує подвійний шар ліпідів. Остання являє собою протоний канал, по якому про¬тони можуть повертатись в строму хлоропласта. АДФ і фосфат з‘єднуються з ферментом в його каталітичній частині F1. Два протона, переміщуються по градієнту електрохімічного потенціалу по протонному каналу (F0), з‘єднуються з киснем фосфату, утворюючи воду. Втрата кисню активує фосфатну групу, і вона приєднується до АДФ з утворенням АТФ.
Фермент Н+-АТФ-синтетаза активна, поки транспортуються протони. Протони пересуваються, якщо їх концентрація у внутритилакоїдному просторі більше. На кожні два електрони, пере-даних по електронтранспортному ланцюгу, всередині тилакоїду накопичується 4Н+. При поверненні назад в строму хлоропласта двох протонів синтезується одна молекула АТФ.
Темнова фаза фотосинтезу. С3-шлях фотосинтезу (Цикл Кальвіна).
Продукти світлової фази фотосинтезу АТФ і НАДФ·Н2 використовуються в темновій фазі для відновлення СО2 до рівня вуглеводів. На сьогодні відомі С3- і С4-шляхи фіксації СО2, фотосинтез по типу товстянкових (САМ-метаболізм) і фотодихання.
Шлях відновлення двоокису вуглецю, відкрив американський біохімік М. Кальвін. Цикл Кальвіна відбувається в стромі хлоропласта.
Сумарне рівняння циклу Кальвіна:
3 РуБФ + 3 С02 + 6 НАДФ·Н2 + 9 АТФ →3 РуБФ + ФГА + 3 Н20 + НАДФ+ + 9 АДФ + 8 Фн
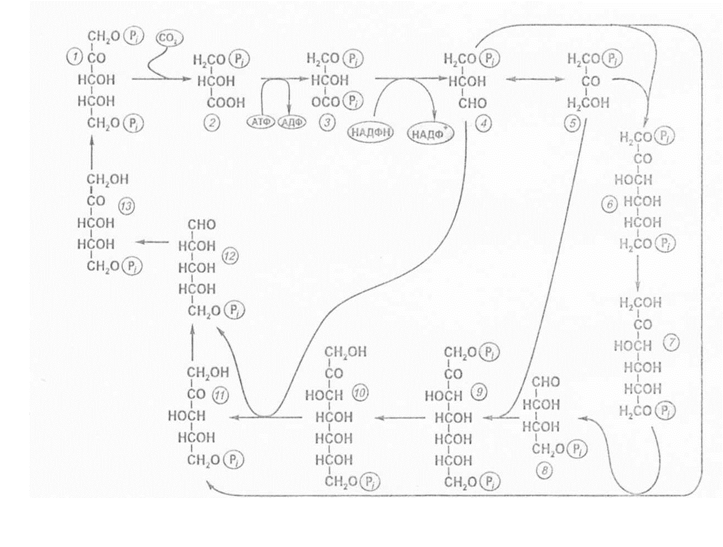 Таким чином, на кожні 3 молекули СО2, що надходять в цикл, утворюється 1 молекула ФГА.
Таким чином, на кожні 3 молекули СО2, що надходять в цикл, утворюється 1 молекула ФГА.
Цикл Кальвіна: 1 – рибулозо-1,5-дифосфат; 2 – фосфогліцеринова кислота; 3 – дифосфогліцеринова кислота; 4 – фосфогліцериновий альдегід; 5 – фосфодіоксиацетон; 6 – фруктозо-1,6-дифосфат; 7 –фруктозо-6-фосфат; 8 – еритрозо-4-фосфат; 9 – седогептулозо-1,7-дифосфат; 10 – седогептулозо-7-фосфат; 11 – ксилулозо-5-фосфат; 12 – рибозо-5-фосфат; 13 – рибулозо-5-фосфат.
Цей цикл поділяють на три етапи: 1) карбоксилювання, 2) відновлення і 3) регенерації.
Перший етап – карбоксилювання рибулозо-бісфосфату і утворення двох молекул фосфогліцеринової кислоти (ФГК). Етап починається з приєднання СО2 до рибулозо-1,5-бісфосфату (РуБФ), який є первинним акцептором СО2. Карбоксилювання катализується рибулозобісфосфаткарбоксилазою (РуБФ-карбоксилазою; РУБІСКА). Продуктом реакції є нестійке шістивуглецеве з'єднання 2-карбокси-3-арабінол-1,5-біфосфат, яке швидко розпадається на дві триози – дві молекули 3-фосфогліцеринової кислоти (3-ФГК).
ФГК є первинним продуктом асиміляції вуглецю. Через те що в її молекулі міститься 3 атома вуглецю, цей цикл отримав іншу назву – «С3-цикл».
Другий етап – відновлення ФГК за допомогою продуктів світлової фази АТФ і НАДФ·Н2 і утворення двох молекул фосфогліцеринового альдегіду (ФГА). Ця реакція єдина реакція відновлення в усьому циклі.
На цьому етапі для подальших перетворень ФГК використовуються продукти світлової фази – АТФ і НАДФ·Н2. Спочатку ФГК фосфорилюється при участі АТФ, і утворюється 1,3-діфосфогліцеринова кислота (ДФГК). Реакція каталізується фосфогліцераткіназою.
Утворена кислота відновлюється за рахунок НАДФ·Н2 і перетворюється на 3-фосфогліцериновий альдегід (ФГА), який частково трансформується за допомогою тріозофосфатізомерази в фосфодіоксиацетон (ФДОА). Процес відновлення ФГК до ФГА каталізується тріозофосфатдегідрогеназою.
Третій етап включає декілька реакцій, в результаті яких утворюється витрачена молекула рибулозо-1,5-бісфосфату. Для цього етапу також потрібна АТФ, що утворилась під час світлової фази фотосинтезу.
В результаті подальших реакцій циклу відбувається регенерація РуБФ. За допомогою альдолази ФДОА з‘єднується з молекулою ФГА, і утворюється молекула фруктозо-1,6-діфосфата (ФДФ).
Фруктозодіфосфат дефосфорилюється і перетворюється на фруктозо-6-фосфат (Ф-6-Ф), що супроводжується накопиченням в стромі хлоропласта неорганічного фосфату. Від фруктозо-6-фосфату відщеплюється двовуглецевий фрагмент (—СО—СН2ОН), який переноситься на третю молекулу ФГА. Реакція відбувається при уча-сті транскетолази. В результаті утворюється пентоза — ксилулозо-5-фосфат, а від Ф-6-Ф залишається 4-вуглецевий цукор – еритрозо-4-фосфат (Е-4-Ф). Еритрозо-4-фосфат з‘єднується з молекулою ФДОА, і утворюється семивуглецевий цукор – седогептулозо-1,7-діфосфат (СДФ). Після відщеплення фосфату седогептулозодифосфат перетворюється в седогептулозофосфат (С-7-Ф). Потім від седогептулозофосфату відщеплюється 2-вуглецевий фрагмент, який переноситься на триозу. Утворюється ще одна молекула ксилулозо-5-фосфат і рибозо-5-фосфат (Р-5-Ф). Обидві молекули ксилулозо-5-фосфат і молекула рибозо-5-фосфат за допомогою рибозофосфатізомерази і рибулозофосфат-3-епімерази перетворюються в три молекули рибулозо-5-фосфат (Ру-5-Ф). Останні фосфорилюються за допомогою АТФ, що утворився на світловій фазі фотосинтезу, і виникають три молекули рибулозо-1,5-бісфосфат (РуБФ). Цю реакцію каталізує фосфорибулокіназа. Таким чином, витрачаються ще три молекули АТФ. Цикл замикається і може розпочатись знову. Рибулозофосфат і ФГА – це кінцеві продукти циклу. Всі інші речовини, що приймають участь, називають проміжними. Оскільки в циклі утворюються пентози, то його ще називають відновлювальним пентозофосфатним циклом (ВПФ-цикл).
Цикл проходить три рази і утворюється 6 молекул ФГА. П’ять з шести молекул тріозофосфату утворюють 3 молекули РуБФ, а шоста молекула ФГА є продуктом асиміляції СО2. Вона може перетворитись в хлоропласті в первинний крохмаль або знову включитись в цикл, або, вийти в цитозоль, бути використаною для утворення сахарози. Із шостої молекули, що залишилася невикористаною 3-ФГА при повторенні циклу під дією альдолази синтезується фруктозо-1,6-діфосфат, з якого можуть утворитися глюкоза, сахароза або крохмаль. Таким чином, для синтезу однієї молекули глюкози (С6) повинно відбутися шість обертів циклу. У кожному оберті циклу використовуються три молекули АТФ (дві для активування двох молекул ФГК й одна при регенерації акцептора СО2 1,5-РДФ) і дві молекули НАДФ·Н2для відновлення ФГК до ФГА. Тому для синтезу однієї молекули глюкози в циклі Кальвіна необхідно 12 НАДФ·Н2 й 18 АТФ. Такий енергетичний внесок світлової фази в темнову фіксацію СО2
Швидкість С3-циклу залежить не тільки від кількості АТФ та НАДФ·Н2, що утворились на світловій фазі, але й від їхнього співвідношення. Тільки співвідношення 3 АТФ до 2 НАДФ+ забезпечує активне відновлення С02 і запасання енергії. Якщо ступінь спряження роботи електрон-транспортного ланцюга фотосинтезу з фотофосфорилюванням незначна, то інтенсивність фотосинтезу знижується через зменшення кількості рибулозо-1,5-бісфосфату, тому що зменшується фосфорилювання рибулозо-5-фосфата і можливість відновлення ФГК до ФГА. В результаті одночасно зі зниженням інтенсивності фотосинтезу серед асимілятів клітини збільшується доля невуглецевих з‘єднань (органічних кислот). Це характерно, для рослин, що вирощуються при низькому освітленні.
С4-шлях фотосинтезу (цикл Хетча-Слека).
Шлях засвоєння СО2, встановлений М. Кальвіним, є основним. Але існує велика група рослин, що включає більше 500 видів вкритонасіннєвих, у яких первинними продуктами фіксації СО2 і відновлення є чотирьохвуглецеві сполуки. Їх називають C4-рослинами. Основи для біохімічного дослідження фотосинтезу в C4-рослин були закладені в роботах Л.А. Незговорової (1956-1957 рр.), Ю.С. Карпілова та І.А. Тарчевского (1960-1963 рр.), а також австралійських вчених Хетча й Слека. В 1966 р. Хетч і Слек запропонували закінчену схему циклу темнових реакцій у C4-рослин, що одержала назву циклу Хетча-Слека.
До C4-рослин належить низка культурних рослин переважно тропічного й субтропічного походження – кукурудза, просо, сорго, цукрова тростина і багато злісних бур'янів – свинорій, просо куряче, просо велике, гумай (сорго алепське), щириця тощо. Як правило, це високопродуктивні рослини, що стійко здійснюють фотосинтез при значних підвищеннях температури й у посушливих умовах.
Для листів C4-рослин характерно анатомічна будова кранц-типу (від німец. Kranz – вінок, корона), тобто наявність яскраво відмінних одна від одної фотосинтезуючих клітин двох типів, які розташовуються концентричними колами: радіально розташовані навколо провідних пучків клітини обкладки й основний мезофіл.
Клітини обкладки провідного пучка містять великі, позбавлені гран (агранальні) хлоропласти. У клітинах мезофілу знаходяться більш дрібні гранальні хлоропласти. Ці два типи клітин фізіологічно не рівноцінні й спеціалізуються на виконанні різних ланок фотосинтезу. C4-цикл можна розділити на дві стадії: карбоксилювання, що відбувається в клітинах мезофілу, та декарбоксилювання і синтез вуглеводів, що відбуваються у клітинах обкладки провідних пучків. Загальним для всіх C4-рослин є те, що карбоксилюванню піддається фосфоенолпіровиноградна кислота (ФЕП) при участі ФЕП-карбоксилази і утворюється щавлевооцетова кислота (ЩОК), що відновлюється до яблучної кислоти або амінуєтся з утворенням аспарагінової кислоти.
СО2, що надходить в рослини в лист крізь продихи, потрапляє в цитоплазму клітин мезофілу, де при участі ФЕП-карбоксилази вступає в реакцію с ФЕП, утворюючи щавлевооцетову кислоту (оксалоацетат). Потім вже в хлоропластах оксалоацетат відновлюється до яблучної кислоти (малату) за рахунок НАДФ·Н2, що утворюється в ході світлової фази фотосинтезу.
Потім малат переноситься в хлоропласти клітин обкладки судинного пучка, де він піддається окисному декарбосилюванню, продуктом якого є піровиноградна кислота (ПВК). Остання знов дифундує в мезофіл, де при участі АТФ, що утворюється в світловій фазі, відбувається регенерація ФЕП, після чого цикл карбоксилювання повторюється за участю нової молекули СО2. СО2 і НАДФ·Н2, що утворюються в результаті окисного декарбоксилювання малату, поступають в цикл Кальвіна, що призводить до утворення ФГК та інших продуктів, властивих С3-рослинам. Тому, саме клітини обкладки виконують роль основної асимілюючої тканини, що постачає цукри в провідну систему. Клітини мезофілу виконують допоміжну функцію – підкачку СО2 для циклу Кальвіна. Таким чином, С4-шлях забезпечує більш повне засвоєння СО2, що особливо важливо для тропічних рослин, де основним лімітуючим фактором фотосинтезу є концентрація СО2. Ефективність засвоєння СО2 С4-рослинами збільшується також за рахунок подачі НАДФ·Н2 в хлоропласти клітин обкладки.
Для С4-цикла характерні наступні особливості: 1) з оксалоацетату – первинного продукту – утворюються малат або аспартат; 2) цикл ділять на два етапи: перший – карбоксилювання ФЕП в клітинах мезофілу, другий – декарбоксилювання малату або аспартату в клітинах обкладки; 3) цикл розділений в просторі: починається і закінчується в основних клітинах мезофілу, а декарбоксилювання відбувається в клітинах обкладки.
С3- і С4-цикли діють сумісно. Таке функціонування двох циклів отримало назву кооперативного фотосинтезу. Основна функція С4-циклу – концентрування СО2 для С3-циклу. С4-цикл є своєрідним насосом (вуглекисневою помпою) для Сз-циклу, за допомогою якого С02 атмосфери переноситься в Сз-цикл.
Якщо С3-цикл є у всіх рослин, то С4-цикл до цього часу виявлений у однодольних та дводольних наземних рослин 16 родин. Рослини, у яких С02 відновлюється в результаті функціонування С3- и С4-циклів, були названі С4-рослинами, а рослини, у яких функціонує тільки Сз-цикл, – С3-рослинами. С4-рослини – це переважно тропічні і субтропічні рослини, вони є високоврожайні.
У С4-рослин більш активними є ФЕП-карбоксилаза, малатдегідрогеназа і аспартатамінотрансфераза, а у С3-рослин – РуБФ-карбоксилаза. ФЕП-карбоксилаза може фіксувати двоокис вуглецю при набагато нижчих її концентраціях, ніж РуБФ-карбоксилаза. Завдяки функціонуванню вуглекисневої помпи (С4-циклу) концентрація С02 в клітинах обкладки, де проходить Сз-цикл, в декілька раз більше, ніж в навколишньому середовищі. В умовах підвищених температур, в яких ростуть С4-рослини розчинність С02 значно знижена. В листках С4-рослин карбокилювання відбувається двічі. Для відновлення ФГК в С3-циклі необхідно АТФ і НАДФ·Н2. Агранальні хлоропласти клітин обкладки містять лише фотосистему I, тому в них можливе тільки циклічне фотофосфорилювання. Тому, в цих клітинах НАДФ не відновлюється. У С4-рослин в хлоропластах клітин обкладки фотосистема I активніша, ніж у С3-рослин, тому у них є циклічний транспорт електронів і пов‘язане з ним фосфорилювання АДФ проходять швидше. Граналь¬ні хлоропласти основних клітин мезофілу містять обидві фотосистеми, тому в них відбувається циклічне і нециклічне фотофосфорилювання з утворенням АТФ і НАДФ·Н2. Коли малат з мезофільних клітин поступає в клітини обкладки і там декарбоксилюється, одночасно відновлюється НАДФ, який необхідний для відновлення ФГК.
Таким чином, С4-цикл постачає в С3-цикл водень для відновлення двоокису вуглецю. Для нормального проходження темнової фази фотосинтезу необхідна жорстка координація швидкостей С3- і С4-циклів і швидкий відтік асимілятів з фотосинтезуючих клітин.
Еволюційно С3-цикл виник раніше, ніж С4-цикл. У деревних, більш старої життєвої форми С4-цикл не виявлений.
Метаболізм органічних кислот по типу товстянкових (САМ-метаболізм).
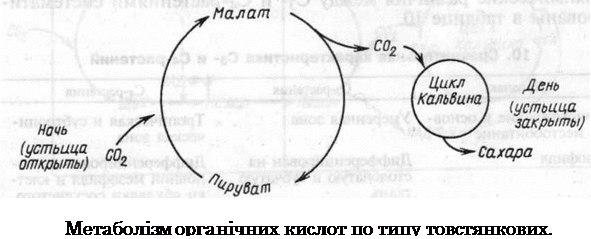 |
Цей тип фотосинтезу вперше був виявлений М.Д. Хетчем у рослин з родини товстянкових (Crassulaceae), тому він отримав назву кислотного метаболізму товстянкових, або англійською Crassulacean acid metabolism (САМ-фотосинтез), а рослини, які його мають – рослини САМ-типу, або САМ-рослини.
Рослини, що відносяться до цього типу ростуть в сухих місцях, тому продихи у них відкриті вночі і зачинені вдень для зменшення транспірації. В даний час цей тип фотосинтезу виявлений у предста-вників 25 родин. Поглинання двоокису вуглецю в темноті було виявлено Н.Т. Соссюром (Швейцарія) ще в 1804 р. СО2, який поглинається, як і у С4-рослин, використовується для карбоксилювання ФЕП. ФЕП утворюється вночі з крохмалю в результаті його гідролізу і включення глюкозо-6-фосфату, що утворюється, в гліколізі. Каталізує реакцію карбоксилювання ФЕП-карбоксилаза. В резуль¬таті утворюється оксалоацетат і ортофосфат:
ФЕП + С02 + Н20 → оксалоацетат + ортофосфат
Оксалоацетат відновлюється до малату за участі малатдегідрогенази:
оксалоацетат + НАДН + Н+ → малат + НАД+
Донором водню є НАДН. Малат накопичується у вакуолі, тому протягом ночі клітинний сік стає більш кислим. На наступний ранок малат виходить з вакуолі в цитозоль. Тут відбувається його окисне декарбоксилювання при участі НАДФ-малатдегідрогенази (декарбоксилюючої), утворюється піруват, С02, і відновлюється НАДФ+:
малат + НАДФ+ → ПВК + С02 + НАДФН + Н+
Піруват фосфорилюється завдяки АТФ при участі піруватфосфатдікінази, і утворюється ФЕП. С02 і НАДФН, які використовуються в С3-циклі.
Крім того, на світлі малат може знову при участі малатдегідрогенази перетворитись в оксалоацетат, який декарбоксилюється під дією ФЕП-карбоксикінази:
малат + НАД+ →оксалоацетат + НАДН + Н+ оксалоацетат + АТФ → ФЕП + АДФ + С02
ФЕП-карбоксилаза, НАДФ-малатдегідрогеназа і ФЕП-карбоксикіназа знаходяться в цитозолі, тому реакції, які вони каталізують відбуваються поза хлоропласту. По мірі використання кислот, що накопичені вночі в вакуолі, величина рН клітинного соку знову збільшується.
ФЕП, який утворився після декарбоксилювання (на світлі) малату або оксалоацетату, в результаті реакцій перетворюється в ФГК або в глюкозо-6-фосфат. Вдень малат інгібує ФЕП-карбоксилазу. ФГК може включитись в цикл Кальвіна в хлоропласті, а глюкозо-6-фосфат – в первинний крохмаль й в такому вигляді накопичуватись, до ночі. Вдень в хлоропластах САМ-рослин відбувається звичайний С3-цикл.
Для цього типу фотосинтезу характерні наступні особливості:
— темнова фаза фотосинтезу розділена у часі: С02 поглинається вночі, а відновлюється вдень. Для сукулентів це дозволяє вдень закривати продихи для зменшення транспірації;
— з оксалоацетату – первинного продукту, як і у С4-рослин, утворюється малат;
— карбоксилювання в тканинах відбувається двічі: вночі карбоксилюється ФЕП, а вдень – РуБФ.
Як і С4-цикл, САМ-тип фотосинтезу є додатковим, він постачає С02 в С3-цикл у рослин, що ростуть в умовах високих температур або нестачі вологи. Вдень (в жару) САМ-рослини запасають СО2, який утворився в результаті дихання, а вночі, коли продихи відкриті, вони поглинають СО2 з повітря. Вночі СО2 дихання зв‘язується ФЕП-карбоксилазою. САМ-фотосинтез допомагає зменшити. транспірацію і зберегти воду, але продуктивність цих рослин набагато нижче, ніж С3-рослин. Співвідношення транспірації до фотосинтезу для САМ-рослин складає 50-100, для С4-рослин — 250-300 і для С3-рослин — 400-500 г транспіруємої води/г поглиненого С02.
У більшості САМ-рослин клітини мезофілу в декілька раз крупніші ніж у С3- й С4-рослин, тому що вони мають дуже великі вакуолі. Кількість продихів у САМ-рослин набагато менша, ніж у С3- й С4-рослин.
У деяких рослин цей цикл функціонує завжди, а у інших — тільки в несприятливих умовах. Перехід к САМ-фотосинтезу відбувається при водному дефіциті і високій температурі вдень та низькій вночі. Перехід до САМ-фотосинтезу може викликати короткий день (деякі види коланхоє) або довгий (деякі види очитка). Деякі САМ-рослини при достатньому зволоженні можуть фотосинтезувати як С3-рослини. Тоді в них продихи відкриваються вдень и закриваються вночі. Іноді у САМ-рослин С02 поглинається і вдень, і вночі.
 |
Фотодихання.
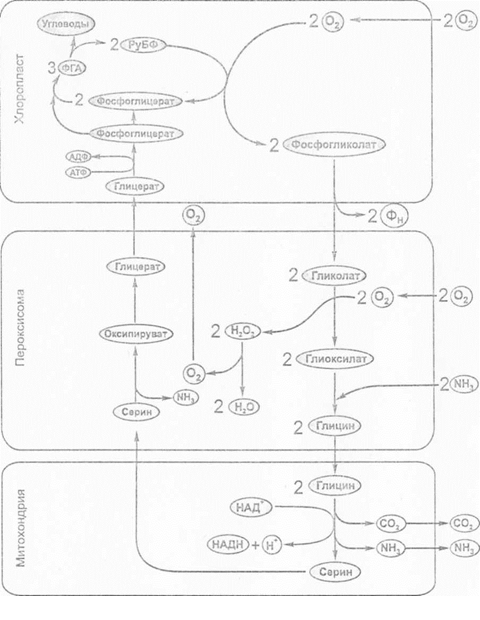
В процесі фотодихання, значну роль відіграє РуБФ-карбоксилаза. При нестачі двоокису вуглецю і великої кількості кисню, цей фермент каталізує приєднання до рибулозо-1,5-бісфосфату не вуглекислого газу, а кисню. Таким чином фермент виконує не тільки карбоксилюючу, але й оксигенеруючу функцію.
Кисень и СО2 є субстратами, що конкурують один з одним за взаємодію з РуБФ-карбоксилазою/оксигеназою (РуБФК/О), таким чином кисень інгібує карбоксилазну, а СО2 - оксигеназну функцію цього ферменту.
Приєднання кисню до молекули РуБФ призводить до того, що замість двох молекул ФГК утворюється одна молекула ФГК і одна молекула фосфогліколата. Фосфогліколат виходить з хлоропласту і при участі фосфогліколатфосфатази дефосфорилюється та перетворюється на гліколат, який поступає в пероксисому. В пероксисомі гліколат реагує з поглиненим киснем; в результаті утворюється гліоксилат і перекис водню. Реакцію каталізує гліколатоксидаза. Перекис розкладається каталазою на воду и кисень, який вивільняється в атмосферу, а гліоксилат в результаті реакції трансамінування при участі гліоксилатгліцинамінотрансферази, перетворюється в гліцин, який транспортується в мітохондрію. В цій органелі з двох молекул гліцину при участі серінгліоксилатамінотрансферази синтезується одна молекула серіну, утворюється СО2, аміак і відновлюється НАД. СО2 вивільняється в атмосферу, тобто втрачається для рослин, або використовується в С3-циклі.
Серин транспортується з мітохондрії в пероксисому, де після дезамінування перетворюється в оксипіруват, який потім відновлюється при участі гліоксилатредуктази в гліцерат за рахунок НАДН. Гліцерат транспортується в хлоропласт, де фосфорилюється за рахунок АТФ в присутності гліцераткінази і утворюється ФГК. ФГК є кінцевим продуктом цього циклу. Вона може включитись в С3-цикл для регенерації РуБФ або використовуватись для синтезу сахарози або крохмалю.
Гліколат є первинним продуктом цього циклу, тому цикл його перетворення в тріозофосфат (ФГК) назвали гліколатним.
Особливості процесу фотодихання:
- С02 утворюється під час перетворення двох молекул гліцина в серін;
- кисень витрачається для синтезу гліколату і гліоксилату. Крім того, якщо НАДН, що утворюється при декарбоксилюванні гліцину в мітохондріях окислюється, то споживання кисню збільшується;
- під час циклу поглинається ки¬сень і вивільняється вуглекислий газ;
- протягом циклу утворюється вільний аміак, який не вивільняється в навколишнє середовище, а використовується для амінування оксиглутарату, в результаті утворюється глутамат;
- ФГК може використовуватись для синтезу сахарози або крохмалю.
Реакції цього циклу відбуваються в хлоропластах, пероксисомах і мітохондріях. В клітині ці органели примикають одна до одної, що підкреслює їхню функціональну залежність.
Протягом гліколатного циклу відбувається втрата СО2, який поглинений в Сз-циклі. Якщо цикл відбувається 10 разів, то з кожних 10 моле¬кул РуБФ, що утворились в Сз-циклі, одна молекула руйнується.
10 РуБФ + 10 О2 → 10 фосфогліколат + 10 ФГК
10 ФГК→ 5ФГК + 5 СО2
10 ФГК + 5 ФГК → 15 ФГК → 9 РуБФ
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 15911; Нарушение авторских прав?; Мы поможем в написании вашей работы!