КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Трахеальные элементы
|
|
|
|
Трахеальные элементы представляют собой наиболее высокоспециализированные клетки ксилемы. Как правило, они вытянуты в длину и в зрелом состоянии мертвы. Для них характерны лигнифицированные оболочки со вторичными утолщениями и порами.
Процесс отложения вторичной оболочки и пропитывание ее лигнином осуществляется еще в живой клетке. При этом на первых этапах развития растения все клетки растущих частей удлиняются (вытягиваются). Однако такое удлинение было бы невозможным при сплошной жесткой оболочке. В этом отношении у высших растений выработалось оптимальное приспособление: вторичная оболочка не одевает клетку сплошь, а расположена кольцами или спиралью. Подобные кольчатые и спиральные утолщения позволяют молодым трахеальным элементам вытягиваться в длину и в то же время препятствуют из сдавливанию. Кроме того, этот тип вторичного утолщения является чрезвычайно экономичным.
При всех достоинствах кольчатых и спиральных элементов как путей для проведения воды, их механическая прочность оставляет желать лучшего. Поэтому, как только у молодого растения заканчиваются ростовые процессы, связанные с удлинением клеток, в ксилеме начинают формироваться трахеальные элементы со сплошной вторичной одревесневшей оболочкой. Когда формирование сплошной оболочки заканчивается, клетки очень быстро отмирают.
Однако и в этом случае оболочка водопроводящего элемента не может быть совершенно сплошной. Обычно в ней имеются многочисленные углубления в виде пор. Поэтому зрелые водопровдящие элементы называют точечно-поровыми. Итак, в процессе онтогенеза (индивидуального развития растения) наблюдается ряд взаимопревращения трахеальных элементов: кольчатые, спиральные, сетчатые, лестничные, точечно-поровые.
Различают два типа проводящих элементов: 1) трахеиды и 2) членики сосудов. Отличаются они главным образом тем, что членики сосудов имеют сквозные отверстия ≈ перфорации, в то время как трахеиды являются неперфорированными элементами. Поэтому по сосудам растворы продвигаются значительно легче, чем по трахеидам.
Трахеиды ≈ это основной водопроводящий элемент высших растений с момента их появления на суше и по сей день. Трахеиды имеют замкнутую со всех сторон первичную оболочку. Поэтому вода по трахеидам должна проходить через окаймленные поры, просачиваясь через первичные оболочки и склеивающий их межклеточный слой. Понятно, что подобная структура не является оптимальной для быстрой подачи воды.
Поэтому в процессе эволюции в ряде групп высших растений возник новый более совершенный проводящий элемент ≈ членик сосуда. Для члеников сосудов характерно наличие в концах клеток перфораций. Соединяясь между собой перфорациями, сотни и тысячи члеников образуют сосуды ≈ длинные сквозные трубки, по которым вода перемещается практически беспрепятственно.
Водопроводящие элементы, обладающие достаточно толстой оболочкой, инкрустированной лигнином, играют в теле растения также и механическую роль.
Поэтому специализация трахеальных элементов в процессе эволюции шла в направлении разделения механической и проводящей функций.
Примитивная древесина большинства хвощей, папоротников, голосеменных имеет гомогенную (однородную) ксилему и состоит исключительно из трахеид и небольшого количества древесинной паренхимы. В такой древесине узкопросветные толстостенные трахеиды выполняют преимущественно механическую функцию, а широкопросветные и тонкостенные ≈ водопроводящую.
Более совершенная гетерогенная древесина покрытосеменных состоит из сосудов, трахеид, волокон - либриформа и запасающей паренхимы.
Членики сосудов у цветковых растений морфологически довольно разнообразны и образуют четкий эволюционный ряд. Этот эволюционный ряд, построенный на работах выдающегося американского ботаника И. Бэйли, помещается во всех учебниках, как одна из ярких и бесспорных демонстраций эволюционных преобразований.
Эволюционный ряд члеников сосудов начинается с длинных трахеид, имеющих лестничную поровость и косые заостренные концы. Постепенно клетки укорачиваются, становятся более широкими, а их концевые стенки все менее наклонными и в конце концов оказываются поперечными.
В наиболее примитивных члениках лестничная перфорационная пластинка имеет множество перегородок. В процессе эволюции они сокращаются и исчезают совсем, так что остается одна сквозная крупная перфорация.
Характерно, что с появлением сосудов цветковых растений не исчезли трахеиды, поскольку совершенный водопроводящий аппарат не всегда является преимуществом. Так, в условиях высокой влажности и затенения способность быстро проводить воду не имеет особого значения. У таких растений процент трахеид и примитивных члеников сосудов с лестничной перфорацией заметно выше, чем у растений, произрастающих в засушливых условиях.
Получается, что соотношение проводящих элементов четко обусловлено экологическими условиями и определяет водный баланс растения.
Мы уже говорили, что сочетание в одном клеточном элементе ≈ трахеиде ≈ и способности проводить воду и механической прочности было важным эволюционным приспособлением. Появление сосудов, с их все увеличивающейся полостью и относительно тонкими стенками, несколько ослабило бы механическую роль ксилемы, если сосуды остались бы единственным элементом этой ткани. В связи с этим наметился второй эволюционный ряд специализации в сторону повышения механической прочности, и трахеиды, через промежуточную стадию волокнистой трахеиды превратились в волокна либриформа.
При этом происходило утолщение оболочек клеток, сужение полостей, все большая редукция окаймления пор. Поры у волокон либриформа стали узкие, щелевидные. Вместе с тем, сократилось и количество пор.
Паренхимные клетки, входящие в состав древесины, запасают крахмал, жиры и многие другие эргастические вещества.
В ходе онтогенеза у растения из первичной латеральной меристемы прокамбия появляется первичная ксилема. У некоторых растений со временем начинает работать вторичная боковая меристема ≈ камбий, дающая начало вторичной ксилеме.
Обычно первичная ксилема отчетливо подразделяется на два структурных типа: 1) протоксилему и 2) метаксилему.
Протоксилема возникает несколько раньше. Это первая стадия развития первичной ксилемы. Обычно протоксилема содержит только трахеальные элементы, погруженные в паренхиму, причем с кольчатыми утолщениями оболочек: в это время клетки еще растягиваются.
Метаксилема более сложная по составу ткань. В ней помимо трахеальных элементов, могут содержаться и волокна, а утолщения оболочек более мощные.
Граница между этими возрастными стадиями проводится весьма приблизительно.
Флоэма (луб)
Флоэма ≈ ткань сосудистых растений, проводящая пластические вещества, образованные в результате процесса фотосинтеза в зеленых клетках листа, от кроны вниз, к корням, а также вверх, к цветам и плодам. Подобно ксилеме, флоэма состоит из нескольких типов клеток и по времени своего возникновения может подразделяться на первичную и вторичную. Первичная флоэма возникает из прокамбия. Вторичная флоэма происходит из камбия.
Первичная и вторичная флоэма содержит одинаковые типы клеток:
ситовидные элементы, которые представлены либо ситовидными клетками, либо члениками ситовидных трубок с клетками спутницами ≈ обеспечивают дальний транспорт пластических веществ;
склеренхимные клетки 2-х типов: волокна и склереиды ≈ несут опорную функцию;
разнообразные паренхимные клетки запасают и транспортируют пластические вещества в радиальном направлении.
Так же как и элементы ксилемы, элементы флоэмы в процессе эволюции высших растений претерпели целый ряд значительных морфологических и функциональных изменений. Однако надо подчеркнуть, что если водопроводящие элементы явились новым приобретением у первых высших растений, осуществивших выход на сушу, то элементы, аналогичные флоэме, уже имелись у некоторых водорослей, и, таким образом, в эволюционном плане флоэма значительно старше, чем ксилема. Более того, можно сказать, что основной физиологический признак клеток флоэмы ≈ их способность проводить растворенные органические вещества ≈ в той или иной степени присущ любой живой клетке. И, следовательно, механизмы, обеспечивающие эту способность, возникли на самых первых этапах развития жизни на Земле. Само собой разумеется, что эта способность была значительно развита и улучшена.
Рассматривая эволюцию флоэмы, нужно отметить, что тонкие целлюлозные оболочки ее клеток несравненно хуже сохраняются в ископаемом состоянии, чем одревесневшие клетки ксилемы. Менее долговечна флоэма и в онтогенезе растения. Обычно новые слои флоэмы достаточно быстро сплющиваются и погибают под напором нарастающих внутренних тканей.
В силу плохой сохранности наши сведения о последовательности этапов эволюции флоэмы гораздо более отрывочны, чем для ксилемы. Но даже по имеющимся данным можно утверждать, что у самых первых наземных растений ≈ риниофитов ≈ имелись клетки вытянутой формы подобные флоэме. Уже эти клетки имели на стенках скопления своеобразно устроенных отверстий, образующих ситечко, или ситовидное поле, по более строгой терминологии.
Итак, межклеточный контакт ситовидных элементов осуществляется при посредстве ситовидных полей.
Ситовидные поля ≈ это участки оболочки, пронизанные многочисленными отверстиями ≈ перфорациями. По сути, это скопление плазмодесм, тончайших тяжей цитоплазмы, пронизывающих клеточную оболочку и связывающих протопласты соседних клеток.
В ситовидных пластинках с крупными отверстиями плазмодесмы сливаются в единый цитоплазменный пояс, образуя гигантскую плазмодесму.
В классическом случае каждое отверстие ситовидного поля облицовано особым углеводом ≈ каллозой, которая покрывает цитоплазменный тяж чехлом. Кроме того, у многих растений каллоза окружает небольшим валиком отверстия канала. Чем старше ситовидный элемент, тем больше в нем накапливается каллозы.
По мере того, как ситовидный элемент переходит в состояние покоя, каллоза закупоривает перфорации и образует подушечку над отверстиями ситовидных полей.
Детальное исследование показало, что: 1) клетки, находящиеся в функционально активном состоянии, содержат очень мало каллозы или не содержат ее вообще; 2) каллоза исключительно быстро синтезируется в ответ на поранения и при окончании функционирования ситовидного элемента.
Различают два типа ситовидных элементов. Более примитивные элементы присущи споровым и голосеменным растениям. Они называются ситовидными клетками. Ситовидные клетки заметно вытянуты в длину и обладают заостренными концами, ситовидные поля рассеяны по боковым стенкам.
Для цветковых растений характерны ситовидные трубки, состоящие из члеников и своеобразных сопровождающих клеток ≈ клеток-спутниц. Членики ситовидных трубок имеют крупные, хорошо выраженные перфорации, располагающиеся на их концах, конечные стенки становятся практически поперечными или слегка скошены.
Таким образом, последовательность эволюционных изменений в ситовидных элементах во многих отношениях сходна с эволюцией члеников сосудов ксилемы. Специализация сопровождалась сокращением числа ситовидных полей, увеличением размеров перфораций и уменьшением угла наклона концевых стенок.
Чрезвычайно важным признаком организации ситовидных элементов является отсутствие в этих клетках клеточного ядра при весьма высокой жизненной активности. Ситовидные элементы в этом отношении представляют собой уникальное явление среди всех остальных клеток. Впрочем, полное отсутствие ядра отмечено только у цветковых растений, тогда как ситовидные элементы голосеменных и других высших растений в большинстве случаев сохраняют ядро, но, как правило, в полуразрушенном состоянии. Такие сморщенные ядра почти всегда лишены ядрышка и называются некротическими.
В противоположность трахеальным ситовидные элементы в зрелом состоянии сохраняют живые протопласты. Внутренняя организация протопласта также не лишена своеобразия. Как правило, в зрелых ситовидных элементах рибосомы полностью отсутствуют, аппарат Гольджи отмечается только на первых стадиях их формирования. Напротив, хорошо развиты митохондрии, пластиды, эндоплазматическая сеть. Вакуоль в этих клетках также не выражена, поэтому цитоплазма находится в разжиженном состоянии.
Ситовидные элементы обычно формируют своеобразные белковые включения, которые прежде назывались "слизистыми тельцами". Теперь их принято называть флоэмным белком или просто Ф-белком. Этот белок имеет вид нитей трубчатой фоэмы со спиральным расположением субъединиц, по своей структуре он напоминает белок мышц животных. Предполагают, что Ф-белок вместе с раневой каллозой участвует в закупорке перфораций при повреждении ткани, предотвращая тем самым вытекание содержимого из ситовидных трубок. Кроме того, высказывается предположение, что Ф-белок, обладающий сократительной реакцией, гонит вещества вдоль по ситовидной клетке.
Отсутствие или полная некротизация ядра в ситовидных элементах находится в разительном противоречии с интенсивной физиологической активностью этих клеток. Поэтому широко распространено мнение, что управление всей жизнедеятельностью ситовидных элементов возложено на ядра соседних клеток.
Так, у цветковых растений членики и связанные с ними клетки-спутники возникают из одной материнской клетки в результате ее митотического деления. Таким образом, эти клетки являются сестринскими и имеют общее происхождение. С каждым ситовидным элементом может быть связана одна или несколько клеток-спутниц.
Предположение о, своего рода, "дистанционном" управлении деятельностью ситовидных элементов со стороны ядра клетки-спутницы широко распространено, но надо признать, что оно не имеет прямого экспериментального подтверждения. Оно скорее основано на убеждении большинства биологов о решающем значении ядра в жизнедеятельности клетки.
Предполагают, что клетки-спутницы регулируют флоэмный транспорт. Кстати и механизм флоэмного транспорта представляет сам по себе большую загадку и сложную проблему. Если движение воды по ксилеме понятно и хорошо объясняется с позиций гидродинамики (вода поднимается по трубкам и капиллярам), то движение веществ по живым клеткам флоэмы невозможно объяснить простой диффузией. Дело в том, что растворы сахаров и аминокислот передвигаются по флоэме со значительно более высокой скоростью (около 1 метра в час).
Для объяснения механизмов флоэмного транспорта выдвигалось множество гипотез. Одна из них ≈ гипотеза тока под давлением, предложенная еще в 1927 г. немецким физиологом Эрнстом Мюнхом, удовлетворительно объясняет экспериментальные данные и в настоящее время поддерживается многими учеными.
Это объяснение является самым простым, поскольку оно основывается исключительно на осмосе, как на движущей силе транспорта ассимилятов. В кратком изложении эта гипотеза утверждает, что ассимиляты транспортируются к месту потребления по градиенту тургорного давления, то есть по градиенту концентрации.
Градиент концентрации создается путем активной загрузки ситовидных трубок сахарозой, при этом вода за счет осмоса свободно поступает в эти клетки. Метаболическая энергия, необходимая для загрузки и разгрузки, поставляется клетками-спутницами.
Следовательно, флоэма является единой системой, состоящей из проводящих элементов вдоль всего пути перемещения сахаров от их источников к местам потребления.
Паренхимные клетки являются постоянными компонентами флоэмы и выполняют типичную запасающую функцию. Кроме того, паренхимные клетки, расположенные рядом с ситовидными элементами, могут участвовать в загрузке и разгрузке проводящих элементов.
Проводящие пучки
Обычно проводящие ткани ≈ ксилема и флоэма ≈ образуют в теле растения тяжи, называемые проводящими пучками. Нередко к проводящим пучкам примыкает механическая ткань-склеренхима, такие комбинированные пучки называются сосудисто-волокнистыми или армированными.
По способности ко вторичному утолщению проводящие пучки делят на открытые и закрытые. В открытых пучках закладывается камбий и они открыты для дальнейшего роста (разрастание в толщину); закрытые пучки, не имеющие камбия, закрыты для вторичного утолщения.
Неполные проводящие пучки содержат или только ксилему или только флоэму. Обычно они очень малы по размерам и встречаются сравнительно редко.
Наиболее распространенными являются полные пучки, включающие в свой состав обе проводящих ткани, и флоэму и ксилему. Полные пучки весьма разнообразны по конструкции.
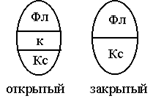 
|  1) в коллатера 1колатеральных пучках флоэма залегает над ксилемой: 1) в коллатера 1колатеральных пучках флоэма залегает над ксилемой:
|
2) биколлатеральные пучки обладают дополнительным солем флоэмы:
3) в концентрических пучках один компонент окружает другой:
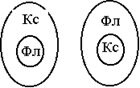 в амфивазальных ксилема окружает флоэму
в амфивазальных ксилема окружает флоэму
в амфикрибральных, наоборот, флоэма окружает ксилему.
|
|
|
|
Дата добавления: 2014-01-06; Просмотров: 1306; Нарушение авторских прав?; Мы поможем в написании вашей работы!