КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Полуцикловые неразрывные характеристики при растяжении. Жесткость материалов при растяжении
|
|
|
|
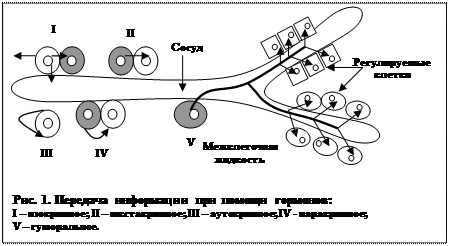
Варианты действия гормонов.
В настоящее время различают следующие варианты действия гормонов:
1) Гормональное, или гемокринное, т.е действие гормона осуществляется на значительном удалении от места образования.
2) Изокринное, или местное, когда химическое вещество, синтезированное в одной клетке, оказывает действие на клетку, расположенную в тесном контакте с первой, а также высвобождение этого вещества осуществляется в межтканевую жидкость и кровь.
3) Нейрокринное, или нейроэндокринное (синаптическое и несинаптическое) действие, когда гормон, высвобождаясь из нервных окончаний, выполняет функцию нейротрансмиттера или нейромодулятора, т.е. вещества, изменяющего (обычно усиливающего) действие нейротрансмиттера.
4) Паракринное – разновидность изокринного действия, но при этом гормон, образующийся в одной клетке, поступает в межклеточную жидкость и влияет на ряд клеток, расположенных в непосредственной близости.
5) Юкстакринное – разновидность паракринного действия, когда гормон не попадает в межклеточную жидкость, а сигнал передается через плазматическую мембрану рядом расположенной другой клетки.
6) Аутокринное действие, когда высвобождающийся из клетки гормон оказывает влияние на ту же клетку, изменяя ее функциональную активность.
7) Солинокринное действие, когда гормон из одной клетки поступает в просвет протока и достигает таким образом другой клетки, оказывая на нее специфическое воздействие (например, некоторые желудочно-кишечные гормоны).
 |
4. Основные этапы образования и превращения гормонов.
В общем виде основные этапы образования гормона следующие:
1) Биосинтез гормона запрограммирован в генетическом аппарате специализированных эндокринных клеток. Он зависит от структуры и экспрессии генов, кодирующих эти гормоны, а также от ферментов, регулирующих синтез гомона. Отсутствие или дефект соответствующих генов приводит к эндокринопатиям, например карликовости при генетическом дефиците гормона роста.
Крупномолекулярные белково-пептидные гормоны синтезируются в полисомах. Затем они передаются в комплекс Гольджи, где «упаковываются» в виде пузырьков – везикул, покрытых тончайшей мембраной. Вместе с гормоном упаковываются АТФ, липиды и ферменты. В таком виде гормон резервируется в эндокринной клетке. Мелкие пептиды, например, тиролиберин гипоталамуса, синтезируется внерибосомальными цитоплазматическими ферментами.
Многие пептидные гормоны (инсулин, кальцитонин и др.) синтезируются в виде предшественника (прекурсора) – прогормона, который затем подвергается энзиматическому расщеплению с освобождением конечной молекулы гормона. Это явление называется прессингом. Так как молекулы прогормона крупнее молекулы дефинитивного гормона, их называют большими (big) гормонами.
Стероидные гормоны и катехоламины образуются с участием цитоплазматических и митохондриальных ферментативных систем. Биосинтез стероидных гормонов происходит из общего предшественника – холестерина. Эти гормоны липофильны, т.е. хорошо растворяются в жирах. В силу этого они содержатся в цитоплазме в липидных каплях и выделяются по мере образования, без предварительного депонирования. Катехоламины синтезируются из аминокислот тирозина и фенилаланина и упаковываются в виде гранул.
2) Секреция и транспорт гормонов.
Секреция гормонов. Гормон, упакованный в везикулы или гранулы, передвигается по направлению к цитоплазматической мембране. Этот процесс осуществляется при участии системы микротубулей или микрофиламентов – субмикроскопиечских трубочек, составляющих клеточный скелет. Для белково-пептидных гормонов и катехоламинов выходу их клетки предшествует взаимодействие цитоплазматической мембраны и мембраны секреторной гранулы, или пузырька, после чего происходит разрушение последних и выброс гормона из клетки. Этот процесс активируется многими факторами – медиаторами, высокой концентрацией калия, электрическими стимулами. Секреция гормона сопровождается затратой энергии, поэтому она всегда сопряжена со сдвигами в системе АТФ-циклический АМФ (цАМФ). Для секреции всегда необходимо участие ионов Са2+, которые активируют белки микротубулярной-микрофиламентозной системы, способствуют взаимодействию с ним гранул гормона, влияют на образование цАМФ. Поэтому снижение содержания Са2+ во внеклеточной среде и поступление его в эндокринную клетку неминуемо приводят к уменьшению секреторной активности этой клетки.
В секреции гормонов, способных к депонированию, различают две фазы:
1 фаза: В ответ на недлительный стимул секретируется ранее синтезированный депонированный гормон.
2 фаза: При длительной стимуляции эндокринной железы выделяется вновь синтезированный гормон.
Стероидные гормоны, содержащиеся в цитоплазме в виде липидных капель, свободно проникают через цитоплазматические мембраны без предшествующего депонирования в секреторных клетках.
Транспорт гормонов. Секретированный гормон попадает во внутреннюю среду организма, преимущественной в кровь, и транспортируется далее.
Большинство гормонов образует в крови комплексные соединения с белками плазмы. Различают специфические транспортные протеины (например, транскортин, связывающий гормоны коры надпочечник) и неспецифические (например, γ-глобулин). Комплексирование (связывание) с белками – обратимый процесс. Связанная и свободная формы могут переходить друг в друга, т.е. между ними устанавливается динамическое равновесие. Часть гормонов в крови находится в связи с форменными элементами крови.
Образование связанной формы гормонов имеет большое физиологическое значение:
1) Это предохраняет организм от избыточного накопления в крови и, следовательно, влияния на ткани свободных гормонов. (Например, при беременности увеличивается продукция гормонов щитовидной железы. Однако основной обмен остается в пределах нормы, так как значительная часть их связывается с белками плазмы. Комплексы гомонов с белком-носителем не проникают через плацентарный барьер, поэтому избыток продукции гормона в материнском организме не оказывает вредного влияния на плод.
2) Связанная форма гормона является его физиологическим резервом. При повышенной потребности организма в определенном гормоне он переходит из связанного состояния в свободное, т.е. увеличивается количество активно действующего гормона.
3) Связывание гормона с белками способствует защите гормона от разрушения ферментами, т.е. продлевает жизнь гормона.
4) Комплексирование с белками препятствует фильтрации мелкомолекулярных гормонов через почечные клубочки, тем самым удерживая эти важные регуляторные продукты в организме.
2) Распознавание гормонального сигнала и его трансдукция (перевод) в биологический ответ.
После поступления в периферические органы гормоны, как правило, отщепляются от белкового компонента и фиксируются в клетках, воспринимающих данный гормон (клетках-мишенях).
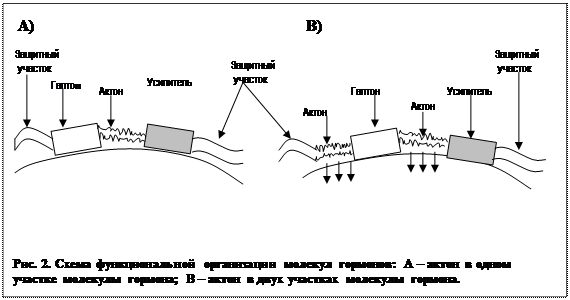
Несмотря на различие в химическом строении, гормоны разных химических групп имеют много общего в организации функциональной группы. В молекуле гормона выделяют - адресный участок, или гаптон, отвечающий за прикрепление всей молекулы к месту специфического гормонального воздействия. Гаптон сам по себе никаким биологическим эффектом не обладает. Другой участок гормональной молекулы – актон, - вызывает физиологические эффекты гормона в клетке-мишени. Сам по себе без гаптона актон к регулируемой клетке не присоединяется. В состав гормональной молекулы входит ряд участков – акцессорных участков - защищающих эту молекулу от воздействия разрушающих ферментов, усиливающих или ослабляющих гормональный эффект. Все указанные участки молекулы могут быть расположены в различных участках гормональной молекулы, а могут и перекрывать друг друга (рис. 1).
 |
Взаимодействие гормона с определенными компонентами клетки называют рецепцией гормона, а клеточные компоненты, взаимодействующие с гормоном, - рецепторами. При взаимодействии гормона с рецептором образуется гормон-рецепторный комплекс, который и воздействует на работу клетки.
Рецепторы могут располагаться внутриклеточно или на поверхности цитоплазматической мембраны. Внутриклеточные рецепторы связывают стероидные и тиреоидные гормоны, мембранные рецепторы – белковые, пептидные гормоны и катехоламины.
В зависимости от того, где образуется гормон-рецептивный комплекс, различают 2 механизма действия гормонов:
1) Рецепция гормонов осуществляется на поверхности клеточной мембраны. Этот тип рецепции свойственен для белково-пептидных гормонов и катехоламинов и обусловлен рядом причин: 1) гормоны этих групп накапливаются в секретирующих клетках, покрытых тончайшей мембраной; 2) молекулы белково-пептидных гормонов очень велики и плохо растворяются в жирах, составляющих основную часть мембран. Проникнуть внутрь клетки через липидные слои мембраны такие молекулы не могут; 3) действие гормонов развивается быстро – за минуты, но продолжается очень недолго.
При таком виде рецепции гормон необходимо перевести внутрь клетки и усилить сигнал, чтобы достигнуть значительного изменения в клетке.
Механизм. Каждая молекула гормона связывается с одним рецептором на поверхности клетки-мишени (рис. 3 А). Пространственная структура рецептора при этом меняется, и он взаимодействует с молекулой G –белка, также встроенную в 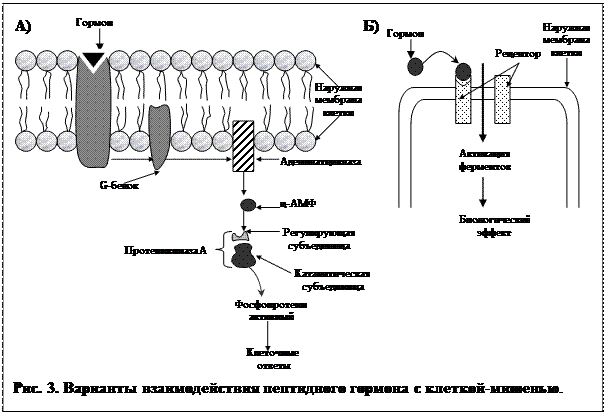 мембрану. G –белок в свою очередь активирует мембранный фермент аденилатциклазу, которая индуцирует у внутренней поверхности мембраны образование большого числа молекул циклического АМФ (ц-АМФ - мощнейший регулятор внутриклеточных процессов). ц-АМФ активирует фермент протеинкиназу А, снимая с нее регулирующую субъединицу. Освободившись от этой субъединицы, протеинкиназа А начинает присоединять к определенным белкам клетки высокоэнергетические фосфатные группы, т.е. фосфорилирует их. Одни из таких белков являются ферментами, другие входят в состав стенок каналов, которые пропускают в клетку или выпускают из нее различные ионы. Фосфорилированные белки резко изменяют свою биологическую активность, и вся работа клетки резко активизируется: мышечная клетка будет сильнее сокращаться, секреторная клетка будет вырабатывать больше веществ. В данном процессе гормон называется первым посредником, ц-АМФ – вторым посредником, протеинкиназа А – внутриклеточным эффектором.
мембрану. G –белок в свою очередь активирует мембранный фермент аденилатциклазу, которая индуцирует у внутренней поверхности мембраны образование большого числа молекул циклического АМФ (ц-АМФ - мощнейший регулятор внутриклеточных процессов). ц-АМФ активирует фермент протеинкиназу А, снимая с нее регулирующую субъединицу. Освободившись от этой субъединицы, протеинкиназа А начинает присоединять к определенным белкам клетки высокоэнергетические фосфатные группы, т.е. фосфорилирует их. Одни из таких белков являются ферментами, другие входят в состав стенок каналов, которые пропускают в клетку или выпускают из нее различные ионы. Фосфорилированные белки резко изменяют свою биологическую активность, и вся работа клетки резко активизируется: мышечная клетка будет сильнее сокращаться, секреторная клетка будет вырабатывать больше веществ. В данном процессе гормон называется первым посредником, ц-АМФ – вторым посредником, протеинкиназа А – внутриклеточным эффектором.
В процессе указанного каскада реакций происходит мощнейшее усиление сигнала. Так, одна молекула АКТГ, взаимодействуя с рецептором менее одной секунды, способна активировать около 1000 молекул аденилатциклазы, которые все вместе способны образовывать более 1 млн. молекул ц-АМФ и т.д. Таким образом, коэффициент усиления в гормональных каскадах способен достигать 107-108. Белково-пептидные гормоны могут и тормозить работу клетки. В этом случае сигнал от рецептора, с которым связался гормон, доходит до особого вида G-белка, который не активирует, а, наоборот, тормозит активность аденилатциклазы.
Некоторые пептидные гормоны, также не проникающие внутрь клетки, имеют другой механизм действия (рис. 3 Б). Связываясь с рецепторами на поверхности клетки-мишени, они открывают в мембране канал, через который в клетку поступают ионы кальция, которые являются активатором целого ряда внутриклеточных процессов, что способствует значительным изменениям деятельности клетки.
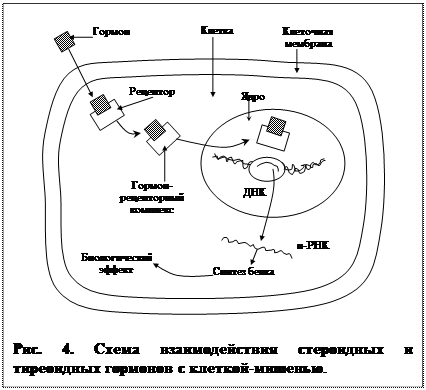 2) Рецепция гормона осуществляется внутри клетки. Этот тип рецепции характерен для стероидов, гормонов щитовидной железы. Эти гормоны жирорастворимы и легко проникают внутрь клетки через наружную мембрану.
2) Рецепция гормона осуществляется внутри клетки. Этот тип рецепции характерен для стероидов, гормонов щитовидной железы. Эти гормоны жирорастворимы и легко проникают внутрь клетки через наружную мембрану.
Механизм. В цитоплазме молекулы гормонов связываются с молекулой рецептора (рис. 4). Комплекс гормон-рецептор проникает внутрь ядра клетки и связывается с участком ДНК, «включая» строго определенные гены.
Идет образование и-РНК, которая выходит из ядра в цитоплазму, где начинается синтез определенного белка или белков, изменяющих деятельность клетки. Эффекты развиваются довольно долго и бывают очень длительными, а часто и программными, изменяя работу клетки на все время ее существования. Именно гормоны с «ядерной рецепцией» ответственны за общую и половую дифференцировку, многие поведенческие изменения. Комплекс гормон-рецептор реагирует с генетическим аппаратом обратимо, и после распада этого комплекса рецепторы могут использоваться многократно.
Одна и та же клетка содержит рецепторы для связывания многих гормонов, в результате чего конечные биологические эффекты являются результирующей многих гормональных влияний. Наряду со специфическим действием, гормоны могут оказывать и другие воздействия. Они могут изменять проницаемость цитоплазматических мембран и мембран субклеточных структур. Например, под влиянием инсулина происходит повышение проницаемости мембран мышечной и жировой ткани к глюкозе; гормоны коры надпочечников, наоборот, стабилизируют мембраны, уменьшая их проницаемость.
Количество гормональных рецепторов в клетке не постоянно. При длительно поддерживающейся высокой концентрации гормона в крови количество свободных рецепторных мест уменьшается (феномен down-регуляции), в результате чего чувствительность клетки (ткани) к гормонам снижается. Наоборот, при сниженном содержании гормона в крови число свободных рецепторов возрастает (феномен up-регуляции). Этот принцип имеет практическое значение. Например, при заболеваниях опорно-двигательного аппарата, сопровождающихся вымыванием кальция из костей, в организм длительно вводят препараты кальцитонина. Вначале эффективны даже малые дозы этого гормона, но затем чувствительность к нему снижается вследствие феномена down-регуляции. В этой связи лечение проводится короткими курсами.
3) Гашение гормонального сигнала имеет большое значение для нормального протекания процессов жизнедеятельности, так как при его задержке возникает опасность избыточного напряжения многих функций.
Механизмы гашения гормонального сигнала:
1) Уменьшение биосинтетической и секреторной активности самих секреторных клеток.
2) Наряду со снижением биосинтеза происходит катаболизм части выделенных гормонов. В результате значительная доля гормонов исчезает из крови вскоре после того, как они были секретированы. Длительность периода полураспада большинства гормонов составляет от нескольких минут до 2 часов; исключение – тиреоидные гормоны (период полураспада значительно дольше). Наиболее полная химическая деградация гормонов происходит в печени и осуществляется ферментами в циклах окислительно-восстановительных реакций путем дезаминирования и метилирования. Как гормоны, так и продукты их катаболизма конъюгируются в печени глюкуроновой и серной кислотами. Это увеличивает их водорастворимость и облегчает последующее выделение через почки. Часть свободных и конъюгированных гормонов поступает в желчь и выводится из организма через кишечник.
3) Эффект down-регуляции рецепторов.
4) Образование антител к гормонам и других ингибиторов, тормозящих биологический эффект.
5) Секреция контррегуляторных гормонов, т.е. гормонов противоположного действия. Например, избыток инсулина вызывает гипогликемию, которая в свою очередь, индуцирует секрецию контринсулярных гормонов.
6) Торможение секреции по принципу отрицательной обратной связи.
К основным полуцикловым неразрывным (характеристикам, получаемым при одноосном растяжении текстильных материалов, относятся:
усилие Ps(t), развиваемое в материале при его растяжении на заданную величину ε за определенное время t;
удлинение материала ε P(t) при действии заданной нагрузки (усилия) Р в течение определенного времени t.
Эти характеристики используют главным образом в исследовательских работах.
Характерная особенность текстильных материалов — их значительная растяжимость. При этом зависимость между нагрузкой и удлинением (рис. 2.9) имеет, как правило, сложный характер, свидетельствующий об изменении жесткости материала по мере его растяжения.
Показатель жесткости выступает как характеристика сопротивления материала, его структурных элементов деформированию. Легкорастяжимые материалы обладают меньшей жесткостью, малорастяжимые — большей жесткостью.
В качестве одной из характеристик жесткости текстильных материалов при растяжении используется модуль жесткости Е (называемый также начальным модулем первого рода, модулем продольной упругости). Модуль жесткости оценивается отношением напряжения σ, развиваемого в материале, к относительной деформации материала ε для участка прямой пропорциональной зависимости на диаграмме напряжение — деформация и выражается в Па.
Модуль жесткости можно также характеризовать углом наклона α 1 и α 2 прямолинейного участка на диаграмме напряжение-деформация (рис. 2.9).
При растяжении упругих материалов модуль жесткости достаточно полно характеризует их жесткость.

Рис. 2.9. Диаграмма напряжение — удлинение материала
Для текстильных материалов модуль жесткости целесообразно оценивать напряжением, вызывающим удлинение материала на 1 %, т. е. напряжением в начальной стадии деформирования, при котором материал сопротивляется изменению размеров и формы.
При удлинении текстильных материалов на 1 % в основном проявляется упругая часть полной деформации материала, а получаемые величины модуля имеют реальный характер.
Проф. А. Н. Соловьев предложил оценивать жесткость материалов при растяжении модулем начальной жесткости Е1, модулем текущей жесткости ЕТ ε и модулем текущей конечной жесткости ЕТ к.

Рис. 2.10. Диаграмма растяжения трикотажа (А. Н. Cоловьева)
Модуль начальной жесткости Е1, Па, соответствует напряжению в образце материала при его растяжении на 1 % и характеризует сопротивление материала деформированию.
Е1= σр / εрк
где σр — напряжение при разрыве, Па; εр — удлинение при разрыве, %; К —показатель жесткости, определяющий характер диаграммы напряжение — удлинение.
Показатель К рассчитывается по формуле
К=(1-η)/ η
где η — коэффициент полноты диаграммы напряжение — удлинение.
Модуль начальной жесткости Е1 достаточно полно характеризует сопротивление деформированию малорастяжимых материалов.
Сопротивление легкорастяжимых материалов модуль E1 характеризует ориентировочно.
По данным проф. А.И.Коблякова, значения модуля E1 для трикотажных полотен очень малы и составляют 1•10-3—1•10-4 мкПа. Причем при испытании полотна по ширине величина Е1 на 2—8 порядков меньше, чем при испытании по длине.
Для легкорастяжимых материалов при расчете модуля начальной жесткости А. Н. Соловьев предложил не учитывать начальную зону диаграммы (рис. 2.10), так как в этой зоне жесткость материала практически не проявляется.
В этом случае начальный модуль жесткости Е z+1, Па, для второй зоны
Е z+1= σр / (εр –z) 
где σр — напряжение при разрыве, Па;
εр — удлинение при разрыве, %
К2 —показатель жесткости, определяющий характер диаграммы напряжение — удлинение во второй зоне.
Где К2 =(1-η2)/ η2
η2=S1/S2
Здесь S1 — площадь фигуры ACD (рис. 2.10);
S2 — площадь фигуры AFCD (точка А — начало отхода кривой растяжения от оси абсцисс).
Зависимость напряжение — удлинение для второй зоны Диаграммы может быть описана как
σр= Е z+1/ (εр –z)
Модуль текущей жесткости ЕТ ε (при z=0) позволяет оценить сопротивление материала деформированию при любой величине удлинения. Модуль ЕТ ε рассчитывается как первая производная от
σр= Е1 ·ε к
ЕТ ε = d σ/ d ε = К Е1 ·ε к-1
Конечную жесткость материала оценивают модулем текущей конечной жесткости ЕТ ε, рассчитанным для момента разрыва пробы материала (при z=0 и ε = εр) по формуле
ЕТ ε = К Е1 · εр к-1
Методы определения этих характеристик текстильных материалов.
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 1247; Нарушение авторских прав?; Мы поможем в написании вашей работы!