КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Рецептор часто взаимодействуют с разными гормонами
|
|
|
|
Различают три типа рецепторов.
1. Рецепторы первого типа являются белками, имеющими одну трансмембранную полипептидную цепь. Это аллостерические ферменты, активный центр которых расположен на внутренней стороне мембраны. Многие из них являются тирозиновыми протеинкиназами. К этому типу принадлежат рецепторы инсулина, ростовых факторов и цитокинов.
Связывание сигнального вещества ведет к димеризации рецептора. При этом происходит активация фермента и фосфорилирование остатков тирозина в ряде белков. В первую очередь фосфорилируется молекула рецептора (автофосфорилирование). С фосфотирозином связывается SН2-домен белка-переносчика сигнала (см. с. 378), функция которого состоит в передаче сигнала внутриклеточным протеинкиназам.
2. Ионные каналы. Эти рецепторы второго типа являются олигомерными мембранными белками, образующими лиганд-активируемый ионный канал. Связывание лиганда ведет к открыванию канала для ионов Na+, К+ или Cl-. По такому механизму осуществляется действие нейромедиаторов, таких, как ацетилхолин (никотиновые рецепторы: Na+- и К+-каналы) и γ-аминомасляная кислота (А-рецептор: Cl--канал).
3. Рецепторы третьего типа, сопряженные с ГТФ- связывающими белками. Полипептидная цепь этих белков включает семь трансмембранных субъединиц. Такие рецепторы передают сигнал с помощью ГТФ-связывающих белков на белки-эффекторы, которые являются сопряженными ферментами или ионными каналами. Функция этих белков заключается в изменении концентрации ионов или вторичных мессенджеров. Таким образом, связывание сигнального вещества с мембранным рецептором влечет за собой один из трех вариантов внутриклеточного ответа: рецепторные тирозинкиназы активируют внутриклеточные протеинкиназы, активация лиганд-активируемых ионных каналов ведет к изменению концентрации ионов и активация рецепторов, сопряженных с ГТФ-связывающими белками, индуцирует синтез веществ-посредников, вторичных мессенджеров. Все три системы передачи сигнала взаимосвязаны. Так, например, образование вторичного мессенджера цАМФ (сАМР) (см. с. 374) приводит к активации протеинкиназ А [ПК-А (PK-A)], вторичный мессенджер диацилглицерин [ДАГ (DAG)] активирует [ПК-С (PK-C)], а вторичный мессенджер инозит-1,4,5-трифосфат [ИФ3 (InsP3)] вызывает повышение концентрации ионов Са2+ в цитоплазме клетки.
Оба гормона задней доли гипофиза - антидиуретический гормон, регулирующий транспорт Na+ и воды в почках, и окситоцин, действующий на гладкие мышцы сосудоы и матки,- являются октапептидами близкой структуры. При концентрациях, в 10 раз превышающих физиологические, окситоцин может связываться в мозговом слое почек c рецепторами антидиуретического гормона и тем самым препятствовать потере организмом Na+ и воды. В свою очередь, антидиуретический гормон при высоких концентрациях может активировать рецепторы окситоцина и таким образом влиять на секреторную активность молочной железы, на сократимость матки и просвет кровеносных сосудов. В организме животного концентрации этих гормонов таковы, что антидиуретический гормон может связываться только со "своими" рецепторами, регулируя работу почек и не влияя на матку и сосуды, а окситоцин контролирует работу гладкой мускулатуры, не вмешиваясь в процессы фильтрации, происходящие в почках.
При концентрациях, в десятки раз превышающих физиологические, инсулин может связываться с рецепторами соматомединов и влиять, например, на превращение пролина в оксипролин или усиливать включение сульфата в мукополисахариды, образующие хрящевую ткань, т.е. оказывать эффекты, свойственные соматомединам.
При высоких концентрациях лютеотропный гормон может связываться с рецепторами фолликулостимулирующего гормона, а фолликулостимулирующий - с рецепторами лютеотропного. Во всех случаях физиологический ответ определяется типом рецепторов, с которыми произошло связывание.
2.1.2. Вторичные посредники гормонов
цАМФ синтезируется из АТФ при посредстве фермента аденилатциклазы.
цАМФ - основной вторичный посредник. В течение некоторого времени после открытия цАМФ в 1950 году он считался вторичным месенджером для всех гидрофильных гормонов. Связывание этих гормонов с рецептором увеличивает содержание цАМФ в клетке в результате активации фермента аденилатциклазы, превращающего АТФ (аденозинтрифосфат) цАМФ. цАМФ затем стимулирует второй фермент, протеинкиназу, которая фосфорилирует ряд других белков. Последние обычно являются ферментами, которые существуют в активной и неактивной формах, в зависимости от того, подверглись они фосфорилированию или нет.
Таким образом, гормоны, которые действуют через цАМФ, осуществляют свою биологическую функцию путем фосфорилирования специфических ферментов и посредством этого изменяют (увеличивают или уменьшают) их активность.
Наконец, должен существовать некий механизм удаления внутриклеточного цАМФ, после того как гормон прекращает свое существование. Это удаление осуществляется вездесущим ферментом, фосфодиэстеразой, которая превращает цАМФ в неактивный метаболит аденозинмонофосфат (АМФ).
В ряде случаев внеклеточные лиганды после взаимодействия с рецепторами индуцируют образование вторичных мессенджеров через участие GTP-связывающих и GTP-гидролизующих гетеродимерных белков, названных G-белками.
(Преобразование сигнала G-белками
G-белки переносят сигнал с рецептора третьего типа на белки-эффекторы. Они построены из трех субъединиц: α, β и γ. α-cубъединица обладает свойством связывать гуаниновые нуклеотиды [ГТФ (GTP) или ГДФ (GDP)]. Белок проявляет слабую ГТФ-азную активность и похож на другие ГТФ-связывающие белки, такие, как ras (см. с. 384) и фактор элонгации Tu (EF-Tu) (см. с. 248). В неактивном состоянии G-белок связан с ГДФ.
При связывании сигнального вещества с рецептором третьего типа конформация последнего изменяется таким образом, что комплекс приобретает способность связывать G-белок. Ассоциация G-белка с рецептором приводит к обмену ГДФ на ГТФ (1). При этом происходит активация G-белка, он отделяется от рецептора и диссоциирует на α-субъединицу и β,γ-комплекс. ΓΤΦ-α субъединица связывается с белками-эффекторами и изменяет их активность, в результате чего происходит открывание или закрывание ионных каналов, активация или ингибирование ферментов (2). Медленный гидролиз связанного ГТФ до ГДФ переводит α-субъединицу в неактивное состояние и она вновь ассоциирует с β,γ-комплексом, т.е. G-белок возвращается в исходное состояние.)
Внеклеточный лиганд специфически распознается трансмембранным рецептором, который, в свою очередь, активирует соответствующий G-белок, локализованный на цитоплазматической поверхности мембраны. Активированный G-белок изменяет активность эффектора (фермента или белка ионного канала, в данном случае - аденилатциклазы), который повышает внутриклеточную концентрацию вторичного мессенджера (в данном примере - cAMP). Каждый вид рецептора взаимодействует только с определенным представителем семейства G-белков, а каждый G-белок - со специфическим классом эффекторных молекул. Таким образом, в одном конкретном случае гормон или нейромедиатор, реагируя со своим рецептором, вызывает активацию GS-белка, стимулирующего аденилатциклазу. Этот фермент-эффектор превращает внутриклеточный ATP в cAMP - классический вторичный мессенджер. Внутриклеточный уровень cAMP может специфически понижаться под действием фосфодиэстеразы, которая превращает cAMP в 5'-AMP.
cAMP активирует множество cAMP-зависимых протеинкиназ, каждая из которых фосфорилирует определенные белки-субстраты. В большинстве клеток животных присутствуют две cAMP-зависимые протеинкиназы, фосфорилирующие белки-мишени по остаткам Ser и Thr (серин/треониновые киназы).
Специфичность регуляторных воздействий cAMP обеспечивается наличием в клетках определенных типов только им присущих тканеспецифических белков, являющихся субстратами для A-киназ. Например, клетки печени обогащены фосфорилазой-киназой и гликогенсинтазой, активность которых регулируется избирательным фосфорилированием по cAMP-зависимому механизму, что сопровождается накоплением или освобождением углеводов в гепатоцитах, адипоциты обогащены липазой, фосфорилирование которой по тому же механизму приводит к освобождению жирных кислот, и т.д.
При понижении концентрации гормонов во внеклеточной среде внутриклеточное содержание сАМР быстро уменьшается, так как фосфодиэстераза превращает сАМР в 5'-AMP. Одновременно происходит дефосфорилирование белков-мишеней A-киназ под действием фосфатаз. Активность некоторых фосфатаз также регулируется по cAMP- зависимому механизму. Кроме того, большинство клеток синтезирует белок, названный ингибитором протеинкиназы (PKI), который блокирует активность C-субъединиц A-киназы. Это сопровождается инактивацией соответствующих факторов транскрипции и подавлением экспрессии регулируемых ими генов.
2.2.Механизм действия гидрофобных гормонов
Липофильные гормоны (стероиды), легко проходят через клеточную мембрану, связываются с рецептором, локализованном либо в цитоплазме, либо в ядре клетки.
Стероидные гормоны и тиреоидные гормоны имеют очень ограниченную растворимость в воде и для них физически невозможно существование в виде свободного раствора при их известных концентрациях в плазме. При этих концентрациях они циркулируют только потому, что существует группа плазменных белков, которые связывают эти гормоны, удаляя их из свободного раствора, но сохраняя их в плазме (рис.1-7сер). Некоторые из этих белков являются высоко специализированными, они связывают только определенные гормоны и, по-видимому, не имеют никаких других функций. Другие, наоборот, связывают все гидрофобные гормоны. Степень связывания гормона с белком и распределение гормона между различными гормонсвязывающими белками варьирует у разных гормонов.
Однако, во всех случаях суммарный гормональный пул представлен преимущественно связанной формой; в некоторых случаях только 1% или меньше суммарного гормона остается в несвязанном с белком состоянии. Эти цифры заслуживают внимания, так как существует общее мнение, что только свободный гормон является биологически активной формой гормона. Следует отметить, что гормон, связанный с белком-носителем, находится в динамическом равновесии с пулом свободного гормона, который в свою очередь находится в равновесии с гормоном, связанным с рецепторами в органе-мишени. Тем не менее, поскольку гормон связан с белком плазмы, он не может взаимодействовать со своим рецептором и таким образом является неактивным. Однако, очевидно, что помимо облегчения растворения гидрофобных гормонов в плазме, связывающие белки формируют большой гормональный резерв, который при необходимости может быть быстро использован для пополнения пула свободного гормона. Размеры свободного пула в большей степени, чем общий пул гормона обеспечивают поддержание нормальной эндокринной функции.
Величина свободного пула и, таким образом, доступность гормона рецептору может регулироваться изменениями секреции, метаболизма или белкового связывания. Легко понять, что концентрация гормона в плазме будет находиться в прямой зависимости от скорости его секреции. Другие факторы, влияющие на концентрацию свободного гормона, могут быть трудноуловимы, но они также достаточно важны как с познавательной, так и с клинической точки зрения. Например, величина свободного пула будет иметь тенденцию к увеличению при любых состояниях, снижающих скорость обмена гормона (деградацию). Существуют такие физиологические (беременность) и патофизиологические состояния (болезнь печени), при которых происходят изменения баланса свободного и связанного пулов определенных гормонов. С диагностической точки зрения важно понимать, что используемые методы определения этих гормонов позволяют измерять суммарную их концентрацию, т.е. сумму свободного и связанного с белками гормона. В этих случаях, поэтому, необходимо также проводить некое определение размеров связывания гормона с белком, с тем чтобы определить концентрацию свободного гормона.
Гормоны, поступая в кровь, транспортируются ею к органам и тканям. При этом лишь небольшая часть гормона циркулирует в свободной форме в виде водного раствора. Преобладающая же часть его находится в крови в форме обратимых комплексов с белками плазмы и форменными элементами. В этих комплексах составляющие компоненты сцеплены нековалентными связями. Особое значение имеют транспортные белки плазмы, специфически связывающие те или иные гормоны. К специфически связывающим транспортным белкам от- носятся такие белки, кактранскортин, избирательно связывающий кортикостероиды и прогестины, секс-стероидсвязывающий глобулин, образующий комплексы с андрогенами и эстрогенами,тироксинсвязывающий глобулин, образующий комплексы с тиреоидными гормонами, и т.д. Вступая в комплекс с этими белками, гормоны аккумулируются в кровяном русле и, тем самым, временно выключаются из сферы биологического действия и метаболических превращений, т.е. обратимо инактивируются. Активной становится свободная, несвязанная с белками форма. По-видимому, образование комплексов гормонов со специфическими белками плазмы есть механизм буферирования сдвигов концентрации гормонов и их резервирования в кровяном русле.
Стероидные гормоны - один из главных классов гормональных соединений всех видов позвоночных и многих видов беспозвоночных животных. Они являются регуляторами фундаментальных процессов жизнедеятельности многоклеточного организма - координированного роста, дифференцировки, размножения, адаптации, поведения [Хартман Э., 1972].
Действие стероидных гормонов на клетки-мишени осуществляется, главным образом, на уровне регуляции транскрипции генов. Оно опосредуется образованием комплекса гормона со специфическим регуляторным белком - рецептором, узнающим определенные участки ДНК в генах, регулируемых данным гормоном. Таким образом, рецепторы всех стероидных гормонов - лиганд-зависимые факторы транскрипции. Для них характерно значительное сходство аминокислотных последовательностей, идентичная доменная структура и сходный механизм действия. Вместе с близкими им рецепторами тиреоидных гормонов они объединяются в семейство рецепторов стероидных/тиреоидных гормонов, которое входит в более обширное надсемейство регуляторных белков - ядерных рецепторов [ Mangelsford D.J. et al., 1995 ].
Стероидные гормоны синтезируются из холестерина, в основном, в коре надпочечников, тестикулах, яичниках и плаценте; однако, каждая ткань, продуцирующая стероиды, имеет свой собственный характерный профиль продуктов секреции.
Подобно надпочечникам, половые железы продуцируют довольно много стероидов, но лишь некоторые из них обладают гормональной активностью. Образование этих гормонов строго регулируется с помощью петли обратной связи, включающей в себя гипофиз и гипоталамус. Действие половых гормонов опосредовано ядерными механизмами, подобными тем, которые используются кортикостероидами.
Стероидные соединения плохо растворяются в воде и хорошо в органических растворителях и растительных маслах. Все стероидные гормоны благодаря липофильности относительно легко проникают через плазматические липопротеидные мембраны и поэтому могут свободно секретироваться клетками стероидогенных эндокринных желез и входить внутрь реагирующих клеток.
стероиды характеризуются трехмерной пространственной конфигурацией, особенности которой оказывают существенное влияние на их биологическую активность.
Гормоны этого класса представляют полициклические соединения липидной природы, в основе структуры которых, как правило, лежит циклопентанпергидрофенантреновое (стерановое) ядро, состоящее из конденсированных между собой трех насыщенных шестичленных (А,В,С) и одного насыщенного пятичленного кольца (D).
Структурно стероиды отличаются друг от друга количеством и расположением функциональных групп, степенью насыщенности углеводных связей, длиной боковой цепи, прикрепленной к стероидному ядру, и другими химическими характеристиками. Структуры некоторых физиологически важных стероидных гормонов представлены на рис.2-3.
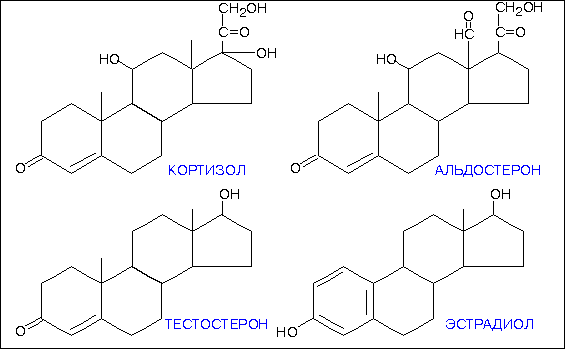
Структура основных стероидных гормонов.
Хотя часто трудно установить определенную взаимосвязь между структурой и активностью, определенные структурные черты, по всей вероятности, являются общими для стероидов со сходной биологической активностью. Например, стероиды с андрогенной или маскулинизирующей активностью, типичным представителем которых является тестостерон, содержат 19 углеродных атомов и не имеют боковой цепи, прикрепленной к стероидному ядру (рис.2-3). Соединения с эстрогенной или феминизирующей активностью, такие как эстрадиол, содержат 18 углеродных атомов и имеют ароматическое кольцо А. Другие физиологически важные стероидные гормоны содержат 21 углеродный атом и включают в себя кортикостероиды и прогестины.
Геометрическая форма стероидной молекулы достаточно стабильна и определяется конформацией колец (циклогексановые кольца имеют конформацию кресла, а циклопентановое - полукресла), характером их соединения (В/С- и С/D-сочленения имеют транс-конфигурацию), а также расположением атомов водорода, радикалов и функциональных групп, присоединенных к стериновому ядру (альфа-или бета-ориентация в пространстве). Следует отметить, что стероиды - оптически активные соединения. Все природные гормоны являются производными D-ряда.
класс стероидных гормонов включает семейства производных четырех углеродных стероидов, содержащих различное количество углеродных атомов (С*4n*5): * прегнана (С*421*5),андростана (С*419*5), эстрана (С*418*5) и холестана (С*427*5).
Прегнановые стероидные гормоны *можно рассматривать как производные С*421*стероида прегнана. Это семейство гормонов состоит из двух основных групп: кортикостероидов - гормонов коры надпочечников и прогестинов *- гормонов яичников и плаценты
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 579; Нарушение авторских прав?; Мы поможем в написании вашей работы!