КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Репликация ДНК
|
|
|
|
Процесс удвоения молекулы ДНК называется репликацией. Репликация обеспечивает точное повторение генетической информации, заключенное в ДНК.
Двойная спираль по форме может быть кольцевой, суперспираллизованной и линейной. В зависимости от формы двойной спирали возможны несколько типов репликации.
Полуконсервативный способ репликации ДНК был проверен в опытах М. Мезельсона и Ф. Сталя (1957 г.). Вся родительская ДНК микроорганизмов содержала ДНК с нуклеотидами в которых был внедрен тяжелый изотоп азота 15N путем выращивания микроорганизмов на среде содержащей 15N. Затем микроорганизмы были перенесены на среду, не содержащую тяжелого изотопа азота. Было установлено, что дочерние клетки микроорганизмов содержали ДНК, состоящей из одной «тяжелой» и одной «легкой» нити. То есть каждая старая нить путем комплиментарной репликации достроила новую нить.
Каким образом может самокопироваться (редуплицироваться) такая стабильная и замкнутая на себя структура, как двойная спираль ДНК? Уотсон и Крик предположили, что ее цепи способны к раскручиванию и последующему частичному разделению вследствие разрыва водородных связей. Образовавшиеся одноцепочечные участки молекулы ДНК могут служить матрицей, к которой на основе принципа комплементарности оснований присоединяются соответствующие нуклеотиды. Между собой эти нуклеотиды связываются фосфодиэфирными связями с образованием новой цепи ДНК. Так как этот процесс происходит на каждой из цепей исходной молекулы ДНК, то в результате образуются 2 двухцепочечные структуры идентичные материнской молекуле. Такой способ назван полуконсервативным, поскольку в каждой вновь образованной молекуле ДНК одна цепь старая (материнская), а другая новая. При делении клетки каждая дочерняя клетка получает такую гибридную молекулу.
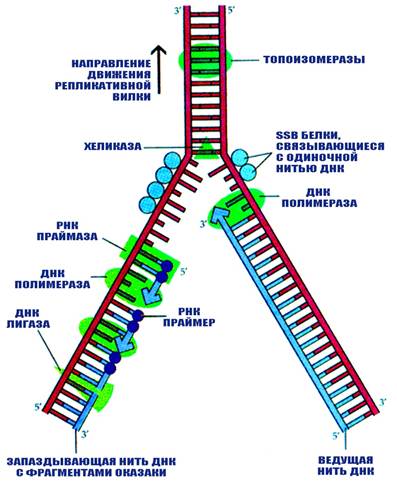
Процесс удвоения молекулы ДНК назван репликацией. В репликации ДНК участвует комплекс ферментов названный реплисомой. В него входят ферменты ДНК-полимеразы, праймазы, топоизомеразы, хеликазы, гиразы, SSB белки, лигазы.
ДНК-полимеразы, или ДНК-зависимые ДНК-полимеразы, это ферменты осуществляющие синтез новой молекулы ДНК на матрице материнской цепи ДНК. Известно несколько типов ДНК-полимераз как у про- так и у эукариот. ДНК-полимеразы осуществляют синтез молекулы ДНК в направлении 5'- 3'. В то же время они обладают экзонуклеазной активностью (то есть способны удалять концевые нуклеотиды) в направлении 3'-5'.
Праймаза синтезирует короткий фрагмент РНК, называемый праймером, комплементарный одноцепочечной матрице ДНК. Праймаза играет ключевую роль в репликации ДНК, так как неизвестно ни одной ДНК-полимеразы, способной начать синтез ДНК без затравки (праймера).
Топоизомераза снимает суперспирализацию ДНК. Топоизомераза I оборачивается вокруг ДНК и вносит разрыв, который позволяет спирали ДНК вращаться и снимает напряжение. После релаксации топоизомераза соединяет разорванные концы.
Гираза также обеспечивает раскручивание суперспиралей ДНК.
Хеликаза перемещается по сахаро-фосфатному остову нуклеиновых кислот и разрывает водородные связи между комплементарно соединенными основаниями.
SSB – белки связываются с однонитевой ДНК предотвращая ее воссоединение с комплементарной цепью или с самой собой с образованием шпилек.
Лигазы осуществляют ковалентное сшивание цепей ДНК в дуплексе. Они образуют фосфодиэфирные мостики между 5'-фосфорильной и 3'-гидроксильной группами соседних дезоксинуклеотидов в местах разрыва ДНК.
Раскручивание и расхождение материнской цепи ДНК происходит в определенных участках, которые названы точками Ori (origin - начало репликации). У прокариот обычно одна точка Ori, а у эукариот множество. Это А-Т богатый участок ДНК, соответственно он легче поддается разделению.
У прокариот имеется одна кольцевая молекула ДНК. При разъединении дуплекса ДНК образуется репликативная вилка, которая может двигаться как в одном, так и в обоих направлениях.
Продвижение ДНК-полимеразы вдоль «матрицы» материнской нити всегда идет в направлении от 3'-конца к 5'-концу. Отсюда следует, что синтезируемая по этой матрице, «комплементарная» к ней новая нить ДНК будет начинаться своим 5'-концом и наращиваться в направлении своего будущего 3'-конца.
Синтез цепей ДНК в обратном направлении не происходит никогда, поэтому синтезируемые цепи в каждой репликативной вилке должны расти в противоположных направлениях. Синтез одной цепи (ведущей, лидирующей) происходит непрерывно, а другой (отстающей) импульсами. Такой механизм репликации называется полунепрерывным. Ведущая цепь растёт от 5¢- к 3¢-концу в направлении движения репликативной вилки и нуждается только в одном акте инициации. Рост отстающей цепи также идёт от 5¢- к 3¢-концу, но в направлении противоположном движению репликативной вилки. Для синтеза отстающей цепи должно произойти несколько актов инициации, в результате чего образуется множество коротких цепей, называемых фрагменты Оказаки в честь открывшего их учёного. Размеры их: 1000-2000 нуклеотидов у прокариот, 100-200 нуклеотидов у эукариот.
По мере движения репликативной вилки концы соседних фрагментов Оказаки соединяются с образованием непрерывной отстающей цепи. Механизмы инициации репликации в точке ori и при образовании фрагментов Оказаки в принципе аналогичны. В обоих случаях происходит образование РНК-затравок (длиной 10-12 нуклеотидов), комплементарных матричной ДНК, в виде продолжения которых синтезируется новая цепь ДНК. В дальнейшем короткие вставки РНК замещаются сегментами ДНК, которые затем объединяются с образованием непрерывных цепей.
Существуют несколько типов репликации. Если молекула ДНК кольцевая, например, как у кишечной палочки (Е. соli) то она начинает удваиваться всегда в одном, строго определенном месте кольцевой ДНК, репликация идет по одной нити в одном направлении и по другой нити в противоположном направлении, пока не встретятся. Затем, под действием релаксирующего фермента, оба конца соединяются. После завершения репликации два полностью синтезированных двуцепочечных кольца разделяются.
Репликация ДНК у эукариотических клеток начинается одновременно во многих точках, которых может быть тысячи. В этих точках репликация идет по одной нити в одном направлении и по другой нити в противоположном направлении. Репликация эукариот идет быстрее чем, у прокариот. Например, в клетках млекопитающих скорость репликации составляет около 60 оснований в секунду.
Полуконсервативная репликация заключается в том, что каждая полинуклеотидная цепь родительской двухцепочечной ДНК является матрицей для синтеза новой комплементарной цепи и две цепи (исходная и вновь образованная), соединяясь, дают следующее поколение ДНК.
Другой тип репликации ДНК наблюдается у некоторых плазмид, репликация идет в одном направлении. Механизм такой репликации ДНК похож на первый, за исключением того, что репликационная вилка движется только в одном направлении.
ДНК в хромомсомах эукариот находится в линейной форме, ее репликация происходит полуконсервативным способом, как предполагали Уотсон и Крик.
Известно, что двухцепочечная спираль ДНК плотно скручена и для ее репликации необходимо, чтобы цепи были разделены хотя бы на короткое время на некоторых участках молекулы. Разделение цепи легко происходит при разрыве водородных связей, соединяющих спаренные основания путем нагревания или действием кислоты или щелочи. В живой клетке разделение двойной спирали осуществляется при участии различных ферментов и других белков. При этом, чтобы произошла репликация разделенные цепи какое-то время должны находиться на некотором расстоянии друг от друга. Эту функцию, и функцию разделения цепей выполняет АТФ-зависимый комплекс ферментов, названный хеликазой или белок rep. Хеликаза разделяет короткие участки ДНК, находящиеся непосредственно перед репликационной вилкой. Участок, где происходит одновременное расплетание родительской ДНК и процесс синтеза, называется репликационной вилкой. Для расплетания двухцепочечной спирали под действием хеликазы требуется энергия, которая выделяется при гидролизе АТФ на АДФ и фосфат. На разделении каждой пары оснований затрачивается примерно две молекулы АТФ. Каждая из разделенных цепей родительской ДНК соединяется прочно с несколькими молекулами белка, который называется ДНК-связывающим белком, который препятствует образованию комплементарных пар и обратному восстановлению цепей. Благодаря этому, разделенные участки ДНК могут функционировать в качестве матрицы для репликации.
|
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 2078; Нарушение авторских прав?; Мы поможем в написании вашей работы!