КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Глубина перемешивания и точка компенсации
|
|
|
|
Предполагая, что в целом скорость фотосинтеза падает по мере уменьшения с глубиной интенсивности света, морские биологи разработали понятие точки компенсации. Благодаря тому что скорость дыхания растительной клетки сравнительно постоянна и не зависит от того, продуцирует растение новые клетки или нет, на кривой выделения кислорода должна существовать точка, в которой расход кислорода при дыхании равен образованию его в процессе фотосинтеза. Это точка компенсации, а глубина, на которой она находится, называется глубиной компенсации. На большей глубине в растительных клетках все еще может происходить фотосинтез, но масса клеток все время сокращается, состояние «здоровья» постепенно ухудшается, а вместе с этим снижается и их способность к размножению или к запасанию жира, благодаря которой они сохраняют плавучесть (рис. 14.5,о).
Однако при активном вертикальном перемешивании точка компенсации может приобретать и другой смысл. Если на минутку отбросить все остальные факторы, планктонные организмы должны проводить по крайней мере столько же времени выше этого уровня, сколько и ниже его. Или же они могут приспособиться «использовать» время, которое они проводят выше точки компенсации, для ускоренного синтеза, а затем, пока организм находится ниже точки компенсации, — просто отдыхать или выполнять другие необходимые для клетки функции, например ассимилировать питательные вещества, чтобы использовать их позднее Понятие «критической» глубины перемешивания.
Глубина компенсации утрачивает свое значение, когда мы имеем дело со всей популяцией фитопланктона. Для этого случая мы вводим другое понятие, которое называется критической глубиной перемешивания. Это глубина, до которой турбулентное перемешивание может распространить всю популяцию, прежде чем «чистая» продукция упадет до нуля. Эта идея (рис. 14.5,6) имеет важные следствия. Если активное перемешивание внутри поверхностного слоя океанских вод происходит до глубин ниже критической, то максимума продукции растений не может быть, так как общая, суммарная потребность в химической энергии для обмена веществ превышает ее производство. Если перемешивание ограничено глубинами меньше критической, имеет место «чистая» продукция, т. е. ее максимум.
* Этим объясняется, почему зимой не бывает никакого «цветения», даже несмотря на то, что зимние штормы перемешивают воду и выносят питательные вещества снизу вверх. «Цветение» начинается только весной, когда с каждым днем увеличивается поддерживающее его солнечное излучение
Очевидно, что это компромиссная ситуация. Перемешивание поддерживает непрерывное возобновление запасов питательных веществ. Чем больше глубина перемешивания, тем выше скорость, с которой поступают новые питательные вещества, и продукция может возрасти, если достаточно света. Но с увеличением глубины перемешивания планктонные организмы больше времени проводят на глубинах ниже их индивидуальных уровней компенсации и общая продукция может снизиться*.
Есть, однако, еще одно условие. Укажем, что чувствительность пигмента, который клетка использует для захвата фотонов, зависит от длины волны. Растение с пигментом, захватывающим фотоны на небольших глубинах, может утратить эффективность поглощения их на более глубоких уровнях из-за того, что длина волны фотона не позволяет ему проникать вглубь, сохраняя достаточную интенсивность. У растений с множеством пигментов, которые покрывают более широкий диапазон поглощаемых длин волн, больше возможностей выжить.
ДОПОЛНИТЕЛЬНЕЙ МАТЕРИАЛ
ИЗМЕНЕНИЕ СОСТАВА СООБЩЕСТВА ФИТОПЛАНКТОНА ВНУТРИ И ВНЕ ОДНОЙ ВОДНОЙ МАССЫ
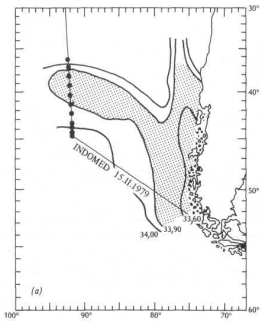
 Общее описание. Из-за удаленности от центров расселения человечества и путей движения юго-восточная часть Тихого океана до сих пор относительно мало изучена. Из имеющихся данных нам известно о поверхностных водах с низкой соленостью, которые простираются на запад от берегов Чили, как видно на рис. 13.5,(7 по изолинии солености 33,9%о. Для этого языка есть два правдоподобных объяснения: 1) выпадающие на море дожди разбавляют поверхностные воды; 2) обильный сток пресной воды с Анд разбавляет поверхностные воды, а течение затем относит их от берега в запад-ком направлении.
Общее описание. Из-за удаленности от центров расселения человечества и путей движения юго-восточная часть Тихого океана до сих пор относительно мало изучена. Из имеющихся данных нам известно о поверхностных водах с низкой соленостью, которые простираются на запад от берегов Чили, как видно на рис. 13.5,(7 по изолинии солености 33,9%о. Для этого языка есть два правдоподобных объяснения: 1) выпадающие на море дожди разбавляют поверхностные воды; 2) обильный сток пресной воды с Анд разбавляет поверхностные воды, а течение затем относит их от берега в запад-ком направлении.
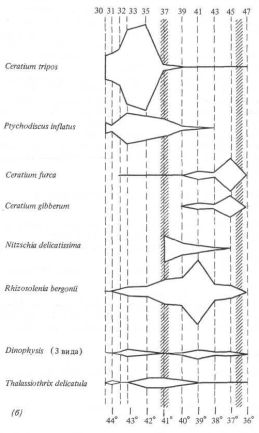
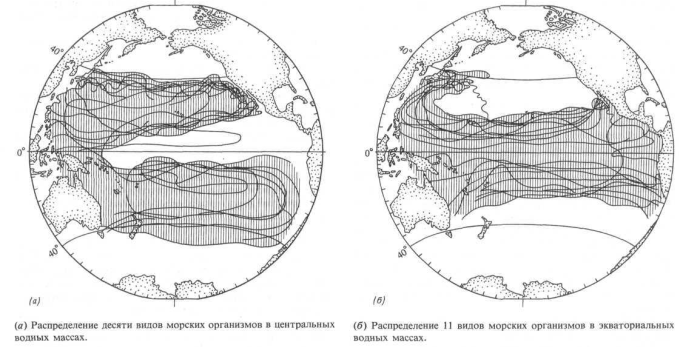
Для проверки второй гипотезы мы выдвигаем логическое предположение, что планктонное «сообщество» внутри языка с низкой соленостью должно существенно отличаться от сообщества, обитающего снаружи. Кроме того, из-за различия водных масс — Центральной воды к северу от языка и Антарктической поверхностной воды к югу от него — внешнее сообщество со стороны экватора должно отличаться от внешнего сообщества', расположенного ближе к полюсу. Во время рейса мы отобрали пробы планктона на 11 станциях, размещение которых показано на рис. 13.5, а. Результаты. Мы определили и оценили количественно все виды планктонных организмов в этих 11 пробах; всего было выделено 26 видов. Каждый из 14 видов составляет более 2% общего количества; основные перечислены на рис. 13.5, б, гае, кроме того, показана относительная плотность популяции на каждой станции наблюдений. Проба 37, взятая у 41
Интерпретация. К югу от места взятия пробы 37 доминирующими видами являются динофлагелляты Ceratium tripos и Ptychodiscus inftatus. В пределах языка с низкой соленостью преобладают диатомей Rhizosolenia hergonii и Nitzschia (два вида). К северу от 37° ю. ш., севернее физической границы этого языка, в планктонном сообществе снова доминируют динофлагелляты, на этот раз Ceratium (три вида).
Вывод. Различие в солености между водами языка с низкой соленостью и внешними водами определенно является биологическим барьером, который препятствует смешиванию видов планктона. Физики до сих пор не доказали, что на этой широте существует западное течение, но приведенные биологические данные отчетливо свидетельствуют о том, что такое течение есть. Dinophysis (3 вида) Thalassiothrix delicamla (6)
на тоже не может служить источником видов-индикаторов просто из-за ее способности мигрировать.
Значит, самые полезные виды-индикаторы следует искать в зоопланктоне. Обычно используют копеподы, хетог-наты и эвфаузииды. Они живут сравнительно долго, их довольно легко поймать, они распространены в широких пределах по вертикали и имеют ограниченную подвижность. Кроме того, довольно хорошо разработана таксономия зоопланктона.
К тому же виды, которым свойственны узкие диапазоны толерантности во время репродуктивно'й фазы, позволяют биогеографам применить дополнительные ограничения к изучаемому сообществу. Некоторые виды определенно должны иметь «свой дом», в котором они размножаются, а это значит, что внутри движущейся водной массы должна происходить циркуляция, к которой вид-индикатор успешно приспособился. Часто эта обстановка представляет собой просто полузамкнутую циркуляцию крупного круговорота, где приток «новой» воды в водную массу в одном месте полностью компенсируется оттоком где-нибудь еще. Зрелые взрослые особи вида-индикатора иногда уносятся из своей биогеографической водной массы. Они могут выжить, но не способны вернуться домой для воспроизведения потомства. Конечно, при таких исследованиях знания о пределах температуры и солености, в которых может происходить размножение, полезны, и получают их при лабораторных экспериментах над самими животными.
Следовательно, по определению биогеографической областью является та, в которой вид-индикатор проходит все фазы развития. Если личиночная стадия очень коротка, мы ожидаем найти одновременно взрослые особи и личинки. Однако если она длится довольно долго, то взрослые особи, как правило, собираются в областях, более благоприятных для питания, и личинки обнаруживаются в другом месте; значит, вся область является биогеографическим «домом».
Водные массы и распределения видов морских организмов
Рис 13.6 Биогеография хетогнат в связи с распределением водных масс.

 Морская биогеография развивается по трем направлениям, которые определяются следующими вопросами:
Морская биогеография развивается по трем направлениям, которые определяются следующими вопросами:
1. Каковы схемы распределения видов и плотности их населения?
2. Благодаря каким факторам существуют наблюдаемые схемы?
3. Что такое сообщество?
Здесь мы опишем распределение видов только для Тихого океана; подобные схемы, но с различными сочетаниями видов типичны для всех крупных океанических бассейнов.
Водные массы
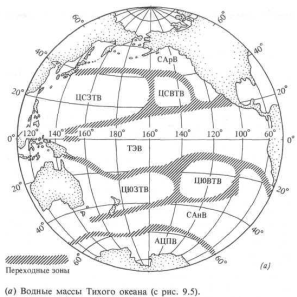
Водные массы Тихого океана схематически показаны на рис. 13.6, а. Они грубо разделены на четыре группы.
Субполярные воды. На севере Тихого океана Субарктическая вода занимает область севернее параллели 45° с. ш.; в южной части Тихого океана Субантарктическая вода расположена к югу от 45° ю. ш. В обоих полушариях субполярные воды движутся по направлению к экватору вдоль восточных границ океанических бассейнов — они отражены в системах Калифорнийского течения и течения Гумбольдта вдоль берегов Северной и Южной Америки.
Ceratium triposPtychodiscus inflatus
Рис 13.7 Биогеорафия птеропод и двух видов копепод относительно водных масс [7].
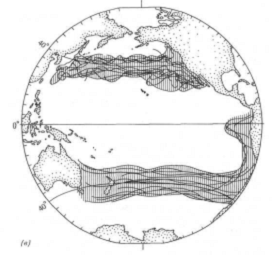 Рис 13.8 Совпадение границ ареалов видов. В целом между границами этих сообществ организмов и границами, определяющими пределы водных масс, существует значительное совпадение.
Рис 13.8 Совпадение границ ареалов видов. В целом между границами этих сообществ организмов и границами, определяющими пределы водных масс, существует значительное совпадение.
Переходные области. Между субполярными и Центральными водами развиты обширные переходные зоны, которые физики долго считали областями «схождения» этих крупных водных масс. В следующем разделе мы увидим, что биологи определяют их как зоны перехода, поскольку вдоль этих границ обнаруживаются разные виды организмов из обеих водных масс.
 Типичные примеры распределения видов
Типичные примеры распределения видов
Субарктические воды. На рис. 13.6, 6 показано распределение в субарктических водах хетогнат вида Sagitta elegans. Эти животные занимают самую северную часть обширного дрейфового течения Западных Ветров, которое мы называем Северо-Тихоокеанским течением, и распространяются понаправлению к полюсу в Берингово море. Если мы сравним карту циркуляции, изображенную на рис. 11.1, с распределением вида 5. elegans, сразу станет ясно, что «дом» этих животных четко совпадает с полузамкнутым Субарктическим круговоротом.
Рис 13.9 Области, занятые сообществами видов Субарктических и Центральных вод, при наложении на T, S – диаграммы для этих регионов океана [5].
Этот круговорот определяется северной ветвью Северо-Тихоокеанского течения, Аляскинским течением, проходящим вдоль берегов Канады и Аляски, потоком Аляскинского течения, идущим вдоль цепи Алеутских островов в Берингово море через проливы между этими островами, и Камчатским течением, которое замыкает круговорот.
Центральные воды. На рис. 13.7, б показано распределение вила Cavolina inflexa — птеропод из типа Mollusca. В северной части Тихого океана эти животные занимают восточный и западный секторы Центральной воды. Кроме того, этот вид встречается в Центральных водах как Северного, так и Южного полушария; для того чтобы описать подобное распределение вида, используется термин «битропич-ность». Многие виды обитают в обеих тропических областях.
Экваториальная вода. Распределение экваториального вида животных приведено на рис. 13.7, в. Это копеподы вида Eucalanus subcrasstis.
Переходные воды. И Субарктические, и Субантарктические водные массы отделяются от Центральных вод зонами с промежуточными характеристиками воды. Такая зона особенно заметно отличается резкими горизонтальными градиентами температуры. Физики-океанологи называют ее Субтропической зоной конвергенции. На рис. 13.7, г показано распределение другого вида копепод — Eucalanus elongatus hyalinus. Эти растительноядные организмы занимают переходную зону как в Южном, так и в Северном полушарии.
Переходные зоны протягиваются в виде широтных полос примерно от 34 до 45° в обоих полушариях. Как видно на рис. 13.7, г, распределение этого вида является битропи-ческим.
Оценки
Основываясь на известных схемах распределения планктона, подобных этим, можно сделать несколько важных обобщений.
1. Границы распределения большинства видов всех так-
сонов имеют одинаковые положение и форму.
Это важный момент. На рис. 13.8 показаны границы площадей, занятых целым рядом животных, обитающих в центральных, экваториальных и переходных зонах. Соответствие поразительно; основные схемы распределения видов в открытом океане повторяются от таксона к таксону и на всех трофических уровнях от первичных продуцентов до хищников, стоящих на вершинах пищевых цепей.
2. Для огромного числа видов в каждой группе границы
всех занятых ими площадей почти совпадают с физическими
границами водных масс.
Однако границы водных масс не обязательно являются неподвижными по отношению к распределению всех видов, обнаруженных в пределах данной водной массы. Мы должны добавить еще три общих вывода.
3. В пределах водной массы ряд видов может встречать-
ся в изобилии, но границы их распространения могут про-
стираться за пределы собственно водной массы.
4. Многие виды прослеживаются в нескольких водных массах. Их называют космополитными.
5. Другие виды обнаружены только в определенных частях некоторых водных масс.
Благодаря каким факторам существуют наблюдаемые биогеографические схемы?
Во-первых, океан, по-видимому, представляет собой гораздо менее сложную область, чем суша. В океане на единицу объема приходится намного меньше экологических ниш.
Во-вторых, физические процессы, благодаря которым сохраняется форма областей обитания видов, вероятно, связаны с процессами, поддерживающими форму водных масс. Мы можем показать это для северной части Тихого океана, используя метод диаграмм температура—соленость (Г—S). На рис. 13.9 характеристики водных масс нанесены на график, как они были установлены на станциях наблюдений Центральных вод в северо-восточной и северо-западной частях Тихого океана (ЦСВТВ и ЦСЗТВ на рис. 13.7, а). Часть области T—S, занятая сообществом Центральных вод, т. е. десятью видами, географические границы распространения которых показаны на рис. 13.8, а, изображена горизонтальной штриховкой. Обратите внимание на четкую корреляцию между водной массой и зоной, занятой одним сообществом. Подобная корреляция очевидна между Т, 5-областями, занятыми субарктическим сообществом (показаны точками), и характеристиками водных масс, полученными на двух станциях в субарктическом регионе. Заметьте, что занятые центральным и субарктическим сообществами области не перекрываются — между ними имеется даже открытое пространство значений температуры — солености. Если бы мы нанесли на диаграмму Т — S зоны, занятые переходными сообществами, они заполнили бы эти пустые пространства.
Исследуем теперь связь между переходными зонами и относящимися к ним видами организмов. В отличие от предыдущего кажется, что к переходным зонам нельзя в такой же степени применить идею о том, что циркуляция внутри океанических круговоротов тесно связана с циклами развития и питания видов от личинки до взрослой особи, эндемичных по отношению к круговым течениям. Ясно, что движение воды в переходных зонах обладает довольно высокой турбулентностью, что вызвано смешением двух водных масс с различными скоростями и направлениями течений. Если в этой зоне линии тока двух водных масс сходятся, она должна быть, кроме того, зоной нисходящего течения (даунвеллинга) или взаимного проникновения двух водных массе через турбулентные завихрения. Если переходная зона связана с расхождением поверхностных ветров, то вдоль фронтальной зоны может происходить сильный апвеллинг. Истинный смысл только что установленного в том, что в переходных зонах происходят совершенно особые процессы перемешивания, а также что длительность этих процессов различна. Как показывают наблюдения (рис. 13.8, в и 13.9), некоторые группы животных приспособили свои жизненные циклы именно к таким физическим условиям. Переходные зоны своеобразны как в физическом, так и в биологическом- отношении.
До сих пор мы говорили о распределении планктонных форм. Если аналогичным образом рассмотреть распределения крупного нектона, то окажется, что многие крупные формы, как и планктонные виды, привязаны к крупномасштабным круговоротам, где их численность наиболее велика, хотя такие животные способны отплывать на большие расстояния. Действительно, многие из них ежегодно покрывают огромные пространства океана, пересекая две водные массы или больше. Почему же многочисленные данные, полученные для такого нектона, столь хорошо согласуются со схемами распределения планктона? Невозможно утверждать, что популяции крупных и могучих животных сохраняются из-за сравнительно медленных течений.
Причиной должно быть то, что виды крупных животных взаимодействуют с видами, находящимися на более низких трофических уровнях, а популяции этих организмов сохраняются благодаря физическим процессам, которые влияют на структуру водных масс. Можно назвать это связью взаимодействия. С точки зрения экологии мы говорим, что данный вид адаптировался к присутствию и поведению других видов и что такая адаптация — неотъемлемая часть общего механизма обратной связи, который контролирует биогеографию жизни в океане.
Заключение
Существует много способов организовать изучение морской экосистемы.
1. При таксономичном подходе подчеркивается, что каждое сообщество организмов состоит из отдельных видов, и каждому из них присвоено особое наименование, признанное во всем мире.
2. Другой способ организовать изучение морской экосистемы — классификация различных способов питания. Об этой схеме многое можно сказать. Благодаря тому что плотность морской воды почти такая же, что и живой протоплазмы, обломки органического вещества остаются в воде во взвешенном состоянии. Вот почему организмы, питающиеся взвесью, обнаружены в столь многих таксонах. Важнейший признак организмов, питающихся взвесью (фильтраторов), — использование слизистых пленок. Это изящный способ захватывать частицы и кусочки органического вещества, и размеры пользующихся этим приемом животных колеблются от микроскопических (радиолярии) до многих сантиметров в длину у других животных.
3. Некоторые биологи-океанологи специально изучают особые, но полные пищевые цепи. Это истинный экосистем-ный метод.
4. Другая схема изучения — классификация организмов, основанная на их подвижности.
5. Наиболее наглядным с точки зрения изучения океана в целом является биогеографический метод. Здесь мы открываем, что сообщества организмов распределены в океане не беспорядочно. Напротив, существует явная корреляция между физическими процессами, формирующими распределение видов, и целыми сообществами. Установлено, что виды обитают в области, где характеристики воды тоже однородны. Более того, такие виды есть во всех таксонах.
Наконец, зададим самый сложный вопрос. Известно, что организмы первых двух трофических уровней рассеяны по всему объему водной массы. Почему же и более крупный нектон сохраняет связь с тем же самым сообществом, если он явно может мигрировать через переходные зоны между водными массами? Тому могут быть три причины. Первая — если учесть, что организмы претерпели эволюцию, то и связи между ними, такие как взаимодействие между хищниками и жертвами, тоже эволюционировали. Приняв это, мы приходим к выводу, что связи в пределах сообщества нерушимы; если же вид мельчайших организмов продолжает развиваться, то развиваются и более крупные. Вторая причина, возможно, тривиальна: ассоциации организмов сохранились просто по привычке.
Третья причина заключается в химических процессах, происходящих в телах более крупных животных. Развиваясь без сложных систем, регулирующих температуру тела, они адаптировались к узкому диапазону характеристик воды. Судя по тому, что они могут пересекать границы водных масс, это не так.
Контрольные вопросы
1. а) В глубоководной экосистеме открытого океана мало синезеленых водорослей. Фитопланктон представлен в основном олотистыми и пиррофитовыми водорослями. Каково их общее название?
б) Биологи относят животных к двум большим группам,
или множествам, которые не являются таксономическими.
К каким?
в) К каким типам беспозвоночных относятся основные
промысловые виды?
г) К какому классу позвоночных относятся главные про-
мысловые виды?
2. Не менее успешно можно классифицировать морских животных по способу их питания. Опишите пять основных способов питания и приведите пример каждого из них.
3. Многие морские животные питаются взвесью. Кратко расскажите, почему этот способ не обнаружен в наземных, неводных системах. Многие морские животные в своих механизмах питания применяют слизистые пленки. Почему этого не делают наземные животные?
4. Что означает термин «сапрофитная пищевая цепь»? Чем этот способ питания отличается — и отличается ли — от нашего собственного?
5. У коров есть зубы, но зачем они нужны копеподам, размеры которых в среднем не превышают 2 мм? Эксперимент по выяснению эффективности питания копепод, проведенный Флемингом, был классическим в том смысле, что он показал возможность полного выедания всех микроскопических растительных клеток. Верно ли это?
6. Большинство видов копепод не производит мужского и женского потомства; они в основном являются гермафродитами. Почему этот способ размножения выгоден для них?
7. Кратко опишите схему организации' морской биосферы, основанную на подвижности организмов.
8. Несомненно, наиболее интересна схема организации морской биосферы, основанная на выделении биогеографических зон. Развейте эту тему, используя два разных подхода.
9. Вопрос 8 можно поставить иначе. Объясните связи между очень большой протяженностью водных масс (гл. 9) и относительно скудным видовым разнообразием в морской биосфере.
10.Если верно, что ключом к пониманию биогеографии океанов является пассивный перенос фитопланктона циркулирующими водами, почему виды фитопланктона не используют в качестве наилучших индикаторов биогеографических провинций и их границ?
11.Развейте мысль о том, что в качестве биогеографических индикаторов, возможно, лучше использовать отмершие морские растения, чем живые.
12. Опишите, приведя хотя бы один пример, как рас-
пределение видов-индикаторов действительно совпадает с
площадью, занятой конкретной водной массой.
13.Что значит термин космополитный по отношению к виду-индикатору? Почему этот вопрос неверен?
14.На основании ваших знаний о циркуляции вод в океанах схематически изобразите границы главных биогеографических провинций в средней части Тихого океана.
15.Напишите краткий очерк о том, что биогеографическая провинция не должна занимать весь океанический круговорот. Например, на рис. 15.13 показано, что сообщества восточных окраин во всех океанических бассейнах организованы сходным образом и отличаются от сообществ других частей круговорота. Позволяет ли космический снимок, приведенный на рис. 14.8, как-нибудь объяснить, каким образом сообщество восточных окраин может образовать отдельную субпровинцию в составе более крупного сообщества круговорота?
Соотношение между распределениями
по вертикали света и первичной продукции
Зная, что количество световой энергии быстро уменьшается с глубиной, мы можем ожидать, что растения лучше всего развиваются на поверхности моря, где свет наиболее интенсивен. Тем не менее мы обнаруживаем, что это не так'. Наблюдениями давно установлено, что больше всего фитопланктона сосредоточено на глубинах примерно от 7 до 17 м. Чем можно объяснить это явное противоречие между потребностями растений и обеспечением их светом?
Объяснение должно включать физический фактор вертикального движения в приповерхностном слое. Вспомните, что конвекция, обусловленная охлаждением и испарением на поверхности океана, вызывает вертикальное перемешивание; дополнительное перемешивание обеспечивают ветровые волны, которые разрушают и создают турбулентное движение в приповерхностных водах. Все движущиеся воды несут с собой мельчайший фитопланктон, так что какое-то вертикальное распределение растительных клеток неизбежно.
С другой стороны, питательные вещества поступают снизу, из-под перемешанного поверхностного слоя. В пределах огромного большинства океанских акваторий, где не происходит апвеллинг, единственный механизм пополнения запасов пищи — это медленная диффузия богатых питательными веществами вод вверх, в поверхностный слой. Турбулентному движению в поверхностном слое способствует вода, активно поступающая снизу. Мы называем этот процесс турбулентной диффузией.
Следовательно, условия, необходимые для возникновения оптимальной продукции растений, должны существовать на каком-то уровне между поверхностью моря, где выше всего интенсивность света, и низом перемешанного слоя, куда поступают питательные вещества. Интервал от 7 до 17 м, в котором мы обнаруживаем самую высокую концентрацию растений, является в среднем компромиссным уровнем, выбранным самими растениями. Вообще при лабораторных экспериментах обнаружилось, что многие растения являются фотоингибиторными, т. е. их продукция снижается, когда они подвергаются воздействию сильного света. Однако замедление развития под действием света свойственно не всем растениям; между видами существует значительный разброс по той интенсивности освещения, при которой каждый вид достигает оптимальной продукции. Например, динофлагелляты, по-видимому, более терпимы к сильной освещенности, чем диатомеи, так что вклад первых в общую продукцию растений гораздо выше в тропических водах. Диатомеи преобладают в более холодных водах высоких широт, где из-за сравнительно меньшей высоты Солнца над горизонтом максимальная интенсивность освещения ниже.
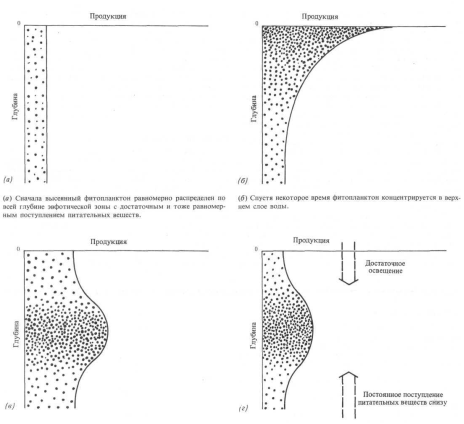
Способность предсказывать первичную продукцию — задача, которую давно пытаются решить биологи-океанологи. Полезным средством для этого является имитационная модель, в которой выделены факторы, предположительно влияющие на поведение растений, а сама модель представляет собой систему математических уравнений, описывающих реакцию растений. Если найдена модель, точно имитирующая реакцию растений в реальном океане, она может работать снова и снова, всякий раз с другим набором весов, приписываемых используемым факторам. Одна такая модель приведена на рис. 14.4. Она начинается с равномерно распределенного планктона, который затем подвергается воздействию изменений в освешенности, обеспечении питательными веществами, выедании и смертности. В результате возникает конечное устойчивое состояние, по которому можно точно предсказать среднее распределение растительных клеток. Такие океанологичекие исследования продолжают проводить, пока еще не разработана универсальная модель.
Глубина перемешивания и точка компенсации
Предполагая, что в целом скорость фотосинтеза падает по мере уменьшения с глубиной интенсивности света, морские биологи разработали понятие точки компенсации. Благодаря тому что скорость дыхания растительной клетки сравнительно постоянна и не зависит от того, продуцирует растение новые клетки или нет, на кривой выделения кислорода должна существовать точка, в которой расход кислорода при дыхании равен образованию его в процессе фотосинтеза. Это точка компенсации, а глубина, на которой она находится, называется глубиной компенсации. На большей глубине в растительных клетках все еще может происходить фотосинтез, но масса клеток все время сокращается, состояние «здоровья» постепенно ухудшается, а вместе с этим снижается и их способность к размножению или к запасанию жира, благодаря которой они сохраняют плавучесть (рис. 14.5,о).
Однако при активном вертикальном перемешивании точка компенсации может приобретать и другой смысл. Если на минутку отбросить все остальные факторы, планктонные организмы должны проводить по крайней мере столько же времени выше этого уровня, сколько и ниже его. Или же они могут приспособиться «использовать» время, которое они проводят выше точки компенсации, для ускоренного синтеза, а затем, пока организм находится ниже точки компенсации, — просто отдыхать или выполнять другие необходимые для клетки функции, например ассимилировать питательные вещества, чтобы использовать их позднее.
Понятие «критической» глубины перемешивания
Глубина компенсации утрачивает свое значение, когда мы имеем дело со всей популяцией фитопланктона. Для этого случая мы вводим другое понятие, которое называется критической глубиной перемешивания. Это глубина, до которой турбулентное перемешивание может распространить всю популяцию, прежде чем «чистая» продукция упадет до нуля. Эта идея (рис. 14.5,6) имеет важные следствия. Если активное перемешивание внутри поверхностного слоя океанских вод происходит до глубин ниже критической, то максимума продукции растений не может быть, так как общая, суммарная потребность в химической энергии для обмена веществ превышает ее производство. Если перемешивание ограничено глубинами меньше критической, имеет место «чистая» продукция, т. е. ее максимум.
Очевидно, что это компромиссная ситуация. Перемешивание поддерживает непрерывное возобновление запасов питательных веществ. Чем больше глубина перемешивания, тем выше скорость, с которой поступают новые питательные вещества, и продукция может возрасти, если достаточно света. Но с увеличением глубины перемешивания планктонные организмы больше времени проводят на глубинах ниже их индивидуальных уровней компенсации и общая продукция может снизиться*.
 Есть, однако, еще одно условие. Укажем, что чувствительность пигмента, который клетка использует для захвата фотонов, зависит от длины волны. Растение с пигментом, захватывающим фотоны на небольших глубинах, может утратить эффективность поглощения их на более глубоких уровнях из-за того, что длина волны фотона не позволяет ему проникать вглубь, сохраняя достаточную интенсивность. У растений с множеством пигментов, которые покрывают более широкий диапазон поглощаемых длин волн, больше возможностей выжить.
Есть, однако, еще одно условие. Укажем, что чувствительность пигмента, который клетка использует для захвата фотонов, зависит от длины волны. Растение с пигментом, захватывающим фотоны на небольших глубинах, может утратить эффективность поглощения их на более глубоких уровнях из-за того, что длина волны фотона не позволяет ему проникать вглубь, сохраняя достаточную интенсивность. У растений с множеством пигментов, которые покрывают более широкий диапазон поглощаемых длин волн, больше возможностей выжить.
* Этим объясняется, почему зимой не бывает никакого «цветения», даже несмотря на то, что зимние штормы перемешивают воду и выносят питательные вещества снизу вверх. «Цветение» начинается только весной, когда с каждым днем увеличивается поддерживающее его солнечное излучение
КАК ИЗМЕРИТЬ ПЕРВИЧНУЮ ПРОДУКЦИЮ В ОКЕАНЕ?
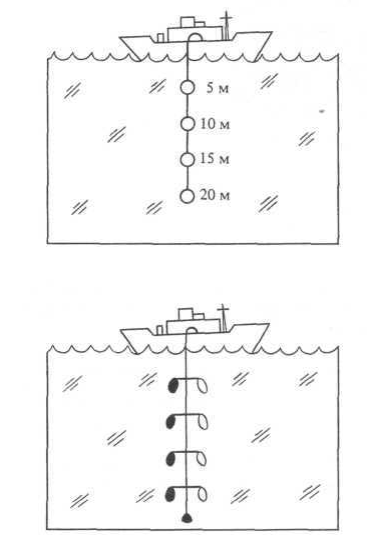
Первичную продукцию следует измерять in situ. Точно воспроизвести реально существующие в океане условия интенсивности освещения и цвета, давления и температуры почти невозможно в лаборатории и еще менее — на борту корабля. В море применяется следующая процедура.
1. Задача — определить первичную продукцию на уровнях 5. 10, 15 и 20 м.
2. Остановив судно, извлекаем пробы воды с каждого желаемого уровня. Фильтруем все пробы, чтобы убрать копепод, поедающих диатомеи; растительные клетки остаются в пробе.
3. Помещаем каждую пробу в три бутыли одинакового объема: одну из прозрачного стекла, пропускающего свет; вторую — черную, чтобы весь свет задерживался; третья бутыль остается на палубе, чтобы измерить в ней содержание кислорода.
4. Опускаем прозрачные и темные бутыли на те же самые уровни глубины, с которых были взяты пробы.
5. По прошествии соответствующего периода времени (не меньше двух часов) возвращаем вес бутыли и тотчас же а) химически фиксируем содержимое прозрачных и темных бутылей, чтобы предупредить дальнейшую физиологическую активность, — иными словами, убиваем пробы, б) измеряем содержание кислорода в каждой бутыли.
6. Разница между содержанием кислорода в прозрачной бутыли плюс количество кислорода в темной бутыли, поглощенного в процессе дыхания, — показатель общей первичной продукции на каждом уровне, с которого были взяты пробы.
Часто используют еще один способ оценки первичной продукции. Он основан на предположении, что содержание хлорофилла в пробе воды пропорционально продукции. Как известно, молекула хлорофилла флуоресцирует. Пробу воды подвергают воздействию вспышки света, чтобы «возбудить» эти молекулы, и затем чувствительный световой приемник считает фотоны, излучаемые молекулами по мере возвращения в нормальное состояние.

Нетрудно оборудовать такой прибор, чтобы проводить непрерывные измерения, пока судно находится в пути, обычно прокачивая морскую воду прямо через флуорометр в судовой лаборатории. Все методы дают значительную погрешность. Поэтому способы измерения первичной продукции продолжают разрабатывать в надежде найти какой нибудь окончательный вариант.
Рис. 14.4. Модель, показывающая, как изменяется с течением времени продукция фитопланктона на разных глубинах в эвфотической зоне. Читателю, вероятно, известно, что модель — это просто система математических уравнений, которые определяют параметры на входе в некоторую систему. Здесь на входе имеются: 1) проникающий свет и доступные питательные вещества; 2) первоначальный запас растительных клеток и 3) скорость реакции планктона на возбуждение. В конце концов модель достигает устойчивого состояния наиболее высокой продукции на уровне, где потребности в свете и пише точно соответствуют существующим условиям.
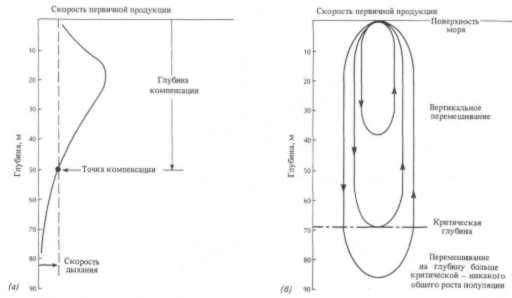
(а) Точка компенсации — это условие, при котором внутри растительной клетки в процессе фотосинтеза образуется столько же кислорода, сколько его расходуется в ходе обмена веществ в организме. Гаубина, на которой это имеет место, зависит от того, как быстро уменьшается с глубиной интенсивность достаточного освещения: таким образом, в чистой воде точка компенсации будет находиться на большей глубине, чем в зеленоватых водах прибрежных областей. Растения могут существовать ниже этих уровней, но они должны расходовать энергию, чтобы поддерживать соответствующее поглощение кислорода.
(б) Критическая глубина относится ко всей популяции фитопланктона и представляет собой глубину, до которой внешние силы — ветер и волны — перемешивают поверхностный слой. Планктонные организмы вблизи поверхности моря в условиях высокой освещенности могут запасать больше энергии химических связей, заключенной в органических веществах, чем они расходуют на обмен веществ. Растительные клетки, попавшие в результате перемешивания на большую глубину, испытывают недостаток освещения и, как правило, производят меньше новой химической энергии, чем расходуют. Когда вертикальное перемешивание ограничено глубинами меньше «критической», вся популяция производит «чистый излишек». Однако перемешивание до большей глубины приводит в результате к «чистому убытку», когда расход в целом превышает продукцию.
Рис 14.4 описание точки компенсации, глубины компенсации и критической глубины.
|
|
|
|
Дата добавления: 2014-01-07; Просмотров: 624; Нарушение авторских прав?; Мы поможем в написании вашей работы!