КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Порушення перетравлення білків та всмоктування амінокислот
|
|
|
|
Перетравлювання білків у тварин з однокамерним шлунком починається у шлунку під впливом ферменту пепсину, який виділяється головними клітинами слизової оболонки у вигляді неактивного проферменту – пепсиногену. Під впливом соляної кислоти шлункового соку пепсиноген унаслідок відщеплення від нього інгібітора перетворюється у пепсин. Оптимальні умови для дії пепсину (рН 1,5–2,0) створює соляна кислота.
Продуктами гідролізу пепсину є поліпептиди, які побудовані із 4–8 або менше амінокислотних залишків, та невелика кількість вільних амінокислот. Подальший гідроліз пептидів і нерозщеплених білків відбувається в тонкому кишечнику під впливом ферментів трипсину і хімотрипсину, які виробляються підшлунковою залозою. Вони виділяються у вигляді неактивних проферментів – трипсиногену і хімо-трипсиногену. Перетворення трипсиногену в трипсин відбувається під впливом ентеропептидази – ферменту кишкового соку. Активний трипсин каталізує утворення нових порцій трипсину із трипсиногену, тобто в подальшому реакція має аутокаталітичний характер.
Хімотрипсиноген перетворюється в активний фермент хімотрипсин під впливом трипсину. Завдяки взаємодії трипсину і хімотрипсину утворюються поліпептиди, низькомолекулярні пептиди та окремі амінокислоти. Подальший розпад поліпептидів у кишечнику здійснюється під впливом пептидгідролаз (ентеропептидаз).
Послідовний вплив усіх протеолітичних ферментів зумовлює пов-ний гідроліз білків корму з утворенням окремих амінокислот, які всмоктуються із кишечнику в кров. Від’єднання окремих амінокислот від білків відбувається у травному каналі людей і тварин нерівномірно. Першими в кишечнику з’являються у вільному вигляді ароматичні амінокислоти і триптофан, тобто в нормі є проміжок часу між появою вільних амінокислот і їх усмоктуванням, який не повинен перевищувати двох годин.
У телят і поросят у перші дні життя основна кількість поживних речовин перетравлюється у кишечнику, оскільки загальна кислотність сичужного (шлункового) соку низька, а вільна соляна кислота зовсім відсутня. У зв’язку з цим показник рН сичужного соку в перші 24–36 год життя близький до нейтрального (4,5–5,1), тому пепсиноген не активується і не руйнує імуноглобуліни молозива. Вільна соляна кислота в сичужному соці телят з’являється наприкінці першої – на початку другої доби життя, а в поросят – на 25–30-й день.
Основним протеолітичним ферментом сичужного соку є хімозин (ренін), під впливом якого казеїноген молозива перетворюється в казеїн, який випадає в осад у вигляді кальцієвої солі, утворюючи пухкий згусток. Імуноглобуліни сироватки молозива евакуюються в кишечник. Таким чином, ферменти сичуга пристосовані до перетравлення лише білків молозива і молока, рослинні ж білки протягом першого місяця життя телят і поросят не гідролізуються. Протеазна і пептидазна активність мікроорганізмів у рубці дуже низька. З віком вона значно зростає, унаслідок чого протеазна і пептидазна деградація протеїну в рубці телят за період з півтора- до 7-місячного віку збіль-шується утричі.
Активність кишкових протеаз висока, а панкреатичної – низька. У перший день життя трипсиноген не активується, оскільки в молозиві є інгібітор та блокатор трипсину, які блокують його перетравлювальну активність, що дозволяє колостральним імуноглобулінам адсорбуватись на слизовій тонкого кишечнику і всмоктуватись у кров. У корів, хворих на мастит, активність блокатора трипсину є меншою майже удвічі, тому імуноглобуліни частково руйнуються в кишечнику (Безух В.М., 1997; Костина М.О., 1997). У подальшому активність панкреатичної протеази підвищується.
У дорослих жуйних білки в рубці гідролізуються ферментами мікроорганізмів, у сичузі і тонкому кишечнику – ферментами макроорганізму. У рубці розщеплюється 50–80 % азотистих речовин кормів, які використовуються для синтезу мікробного білка. Гідроліз білка складається з двох етапів (рис. 8). Спочатку протеїн корму під впливом протеїназ розщеплюється до поліпептидів, які у свою чергу пептидазами гідролізуються до амінокислот. Оптимальною реакцією для протеїназ вважається рН близько 7,0, а при рН 5,7 вона є удвічі нижчою, для пептидаз – від 5,5 до 7,0.
Частина вільних амінокислот, особливо незамінних, відразу ж використовується мікроорганізмами для синтезу білка, незначна частина всмоктується стінкою рубця, а решта – розпадається. Основною реакцією розпаду є дезамінування під впливом бактеріальних дезаміназ. Кінцеві продукти дезамінування – аміак, вуглекислий газ та коротколанцюгові жирні кислоти. Оптимальною реакцією для дії більшості дезаміназ є рН близько 7,0.
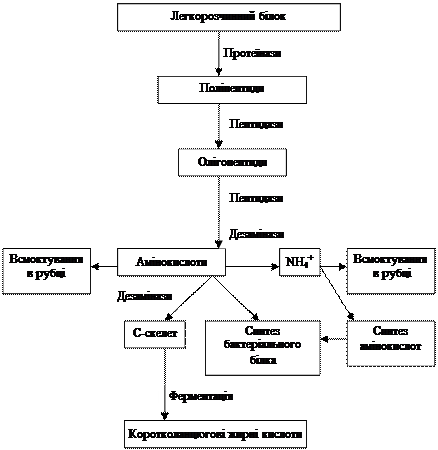 |
Рис. 8. Схема перетворення протеїну в рубці
(Янович В.Г., Сологуб Л.І., 2000)
Аміак, що утворюється в рубці при дезамінуванні амінокислот, частково всмоктується через його стінку у кров, а в основному використовується мікроорганізмами рубця для синтезу амінокислот і білка. Більшість мікроорганізмів рубця (86–92 %) для синтезу білка використовують аміак та амінокислоти, і тільки невелика група бактерій – виключно амінокислоти. Вважається, що лише бактерії здатні використовувати азот аміаку в синтетичних процесах (50–80 % азоту мікроорганізмів походить із азоту аміаку), інфузорії не використовують або ж використовують незначну його кількість. Аміак зв’язується бактеріями двома шляхами – утворенням глутаміну або глутамінової кислоти. Найбільш ефективно використовується аміак у синтезі амінокислот мікроорганізмами при концентрації його в рідині рубця
5 ммоль/л (9 мг/100 мл).
Мікроорганізми рубця в синтезі власних білків використовують синтезовані ними амінокислоти та амінокислоти, які звільняються при розщепленні протеїну кормів. Амінокислоти, що звільняються в результаті розщеплення мікробного протеїну, становлять від 50 до 90 % загальної кількості амінокислот, що всмоктуються в тонкому кишечнику жуйних тварин. Синтез мікробного протеїну в рубці жуйних залежить насамперед від умісту в раціоні енергії і розчинного протеїну. Він становить від 24 до 36 г азоту мікробного протеїну на 1 кг перетравленої в рубці органічної речовини, або 1,0–1,5 г азоту на 1 мДж обмінної енергії. Валовий синтез мікробного протеїну в рубці овець та корів становить відповідно 50–100 і 700–1500 г за добу. Найбільш інтенсивний синтез бактеріального протеїну в рубці корів із добовим надоєм 10–14 кг виявлений при вмісті у сухій речовині раціону 12–12,5 % сирого протеїну, 17–18,5 клітковини і 24–27 % цукру і крохмалю.
Важливим фактором, що впливає на ефективність синтезу бактеріального протеїну в рубці, є ступінь забезпечення потреби мікроорганізмів в енергії, яка коливається в межах від 0,2 до 7,0 ммоль АТФ на 1 г сухої речовини бактеріальної маси за годину. Вважається, що близько 75 % енергії, необхідної для росту мікроорганізмів, витрачається на синтез білка. Цукри і крохмаль найбільшою мірою забезпечують потребу мікроорганізмів в енергії, а високий їхній уміст у раціоні тварин позитивно впливає на ферментативні процеси і синтез бактеріального протеїну в рубці. Проте надлишок крохмалю в раціоні тварин при високому вмісті концентратів призводить до зменшення розщеплювання клітковини, зниження відношення ацетат: пропіонат і збільшення продукції лактату в рубці. Внаслідок цього синтез азоту мікробного білка зменшується до 16–18 г на 1000 г ферментованої в рубці органічної речовини. Зменшення синтезу мікробного білка спостерігається також при високому вмісті в раціоні тварин легкорозщеплюваного протеїну і низькому вмісті цукру та крохмалю. Це має міс-це, зокрема, при згодовуванні тваринам великої кількості силосу.
В останні роки велика увага при нормуванні протеїну високопродуктивним коровам надається співвідношенню між легкорозщеплюваним протеїном раціону і його загальною кількістю. Відомо, що 50–80 % протеїну, який надходить із кормом, у рубці жуйних гідролізується під дією ферментів мікрофлори рубця. Протеїн, що не розщепив-ся в рубці, та білок бактерій і найпростіших надходять у кишечник. При високій розчинності протеїну в рідині рубця, а значить і при швидкому розщепленні його під дією бактеріальних ферментів, утворюється багато аміаку, який рубцева мікрофлора неспроможна повніс-тю використати для синтетичних процесів, що спричинює його великі втрати. Згідно з рекомендаціями, кількість протеїну, що розщеплюється в рубці, повинна становити на початку лактації 50–60 %, а в середині та наприкінці – близько 70 %.
Порушення перетравлення протеїну в передшлунках жуйних спостерігається при ацидозі та алкалозі рубця. При гострому перебігу ацидозу рубця величина рH рідини становить 4,0–5,0, а при хронічному – від 5,0 до 5,8. При кислій реакції (рН 5,7) активність бактеріаль-них протеїназ рубця зменшується удвічі. При ацидозі порушується також перетворення амінокислот. Якщо величина рН знижується до 5 і менше, розщеплення амінокислот може відбуватися під впливом інших бактеріальних ферментів – декарбоксилаз. Оптимальною величиною рН для них є 3,0–5,5. Ці ферменти продукують мікроорганізми групи Coli, клостридії, гриби. При декарбоксилюванні утворюються аміни, більшість з яких є токсичними для тварин. Із орнітину утворюється путресцин, з лізину – кадаверин, гістидину – гістамін, тирозину – тирамін.
Порушення перетравлення білків у шлунку (сичузі) та кишечнику, насамперед, може бути спричинене структурними змінами, які виникають при ураженні цих органів, а також підшлункової залози, при багатьох хворобах (гострому і хронічному гастриті та гастроентериті, панкреатиті, виразковому гастриті та абомазиті, інфекційних та паразитарних хворобах, що супроводжуються ураженням шлунка і кишеч-нику). Так, при рота- і коронавірусному ентеритах новонародженого молодняку віруси руйнують зрілі епітеліальні клітини мікроворсинок тонких кишок, які є структурною основою мембранного травлення, тому знижується синтез ферментів, що завершують гідроліз білків, і змен-шується їхній вихід на поверхню ентероцитів. Зокрема, синтез дипептидаз при тяжкому перебігу зменшується у 30 разів (Щербаков Г.Г., 1987). Унаслідок цього засвоєння протеїну молозива і молока знижується з 98 до 40 %.
При гіпоацидному гастриті зменшується виділення вільної соляної кислоти, а при анацидному – вона відсутня. Відповідно знижується й активність пепсину, що спричинює порушення перетравлення протеїну корму. Хронічний гастрит зі зниженою секрецією НСl часто супроводжується атрофічними процесами у слизовій оболонці шлунка. При цьому зменшується секреція шлункового соку, у ньому відсут-ні соляна кислота і ферменти (ахілія). При гіпоацидному гастриті розслаблюється сфінктер пілоруса, що прискорює евакуацію вмісту шлунка в кишечник; протеїн корму не встигає гідролізуватися до поліпептидів та амінокислот, тобто порушується шлункове, а згодом і кишкове травлення, що спричинює зменшення всмоктування поживних речовин у кишечнику.
Структурні зміни в органах травлення виникають також при хворобах серця, нирок, набряковій хворобі поросят. Унаслідок порушення водного обміну спостерігається набряк слизових оболонок шлунка і кишечнику, порушується їхня секреторна і всмоктувальна функції, знижується активність протеолітичних ферментів. У новонароджених телят-гіпотрофіків слизова оболонка сичуга побудована за ембріональ-ним типом, диференціація ворсинок дванадцятипалої і голодної кишок слабко виражена, що зумовлює зниження протеолітичної активності реніну, пепсину та інших гідролаз.
Порушення перетравлення білків може бути рефлекторним. Спочатку виникають функціональні розлади шлунка і кишок у результаті ураження інших органів: передшлунків, печінки, легень, нирок, підшлункової залози. Функціональні зміни можуть виникати внаслідок посилення екскреції продуктів обміну речовин або токсинів слизовою оболонкою шлунка і кишок при зниженні видільної функції уражених органів (легень, нирок), при порушенні функцій вегетативної нер-вової та ендокринної систем. Тривалі функціональні розлади шлунка і кишечнику рефлекторного і компенсаторного походження здебільшого зумовлюють структурні зміни у їхніх тканинах. Дослідження вчених показали, що при пневмоніях у лошат (Скородумов М.Т.) і поросят (Ковбасенко М.Х.) пригнічується секреторна функція шлунка, а у телят – сичуга, зменшується кількість вільної соляної кислоти, у телят гальмується виділення соку підшлунковою залозою, знижується перетравлювальна активність пепсину та трипсину (Кости-
на М.О.). При гіпотонії та тимпанії рубця секреторна функція сичуга в телят нижча на 50 % (Ганжа І.Д).
Білки, які не піддались перетравленню, та амінокислоти, які не адсорбувались у тонкому кишечнику, надходять у товстий кишечник, де за участі мікроорганізмів розпадаються (гниття білків) з утворенням токсичних амінів (путресцин, кадаверин, гістамін, тирамін) та ароматичних сполук (індол, скатол, крезол), які всмоктуються у кров і діють токсично. Деякі з цих сполук (індол, скатол, крезол, фенол) у печінці знешкоджуються шляхом утворення парних сполук із активними формами сірчаної та глюкуронової кислот. Однією з таких сполук є індоксилсірчана кислота, або індикан, який виділяється із сечею і є показником інтенсивності процесів гниття білків у кишечнику. Індиканурія часто супроводжує недостатню секрецію соляної кислоти у шлунку.
Структурно-функціональні зміни в шлунку і кишечнику спричинюють порушення гідролізу білків та всмоктування амінокислот, подовжують період від утворення окремих амінокислот до їх абсорбції з двох до 5–6-ти годин. Окремі амінокислоти надходять у печінку нерів-номірно, що зменшує синтез білка, особливо при порушенні абсорбції незамінних амінокислот. Інші амінокислоти, що всмоктуються в достатній кількості, використовуються для синтезу білка не повністю, тому їх рештки перетворюються в жири і вуглеводи та виводяться із сечею (аміноацидурія), що поступово знижує ефективність використання кормів, зумовлює гормональні порушення і накопичення в організмі проміжних продуктів обміну амінокислот. Наприклад, зниження засвоєння тирозину зменшує синтез гормонів щитоподібної залози, триптофану – нікотинової кислоти.
Порушення перетравлення білків може проявлятися також кормовою алергією, яка виникає внаслідок того, що частина білків і високомолекулярних пептидів всмоктуються в кишечнику в кров у незміненому вигляді і діють як алергени. У тварин кормова алергія виникає при поїданні кормів, до яких вони не адаптовані (замінники молока із соєю), уражених мікотоксинами, з домішками лікарських препаратів та незвичних для організму хімічних речовин. За незвичайного антигенного кормового навантаження відбувається швидке виснаження механізмів місцевого захисту: при цьому різко знижується вміст у слизовій оболонці Ig A, макрофагоцитів, епітеліолімфоцитів, біфідо- і лактобактерій. У таких умовах відбувається абсорбція антигенів із кишечнику в кров. Унаслідок їхнього контакту з імунокомпетентними клітинами розвивається імунна відповідь і відбувається сенсибілізація організму. У крові збільшується кількість лімфоцитів, еозинофілів та Ig E. Клінічно кормова алергія проявляється ураженням органів шлунково-кишкового каналу: у хворих тварин раптово з’являються абдомінальні болі, нудота, блювання, проноси або запори. З’являються ураження шкіри – набряки, висипи, ділянки еритемного запалення (рис. 9).

Рис. 9. Ураження шкіри у вигляді еритемних плям
при кормовій алергії (За Карпутем І.М.)
|
|
|
|
|
Дата добавления: 2014-01-20; Просмотров: 3258; Нарушение авторских прав?; Мы поможем в написании вашей работы!