КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Реалізація генетичної інформації
|
|
|
|
РОЗДІЛ 5
Лекція 9
План
5.1 Дія генів
5.2 Від поліпептиду до ознаки
5.3 Регуляція генної активності
5.4 Модифікації
5.5 Взаємини алелей
5.6 Полігенне спадкування й плейотропія
Генетична інформація закодована в ДНК. Інформація, що перебуває в клітинному ядрі, являє собою генотип. ДНК, що міститься в одному наборі хромосом, називається геномом, а позаядерна ДНК (у мітохондріях, пластидах й основній речовині цитоплазми) - плазмоном. У бактерій ДНК в еквіваленті ядра являє собою геном, а позаядерна ДНК представлена у формі плазмід. Структури, що складаються із ДНК і зустрічаються в основній речовині цитоплазми, в еукаріот, так само як й у бактерій, називають плазмідами.
Генетична інформація в геномі бактерій і багатьох вірусів укладена в один-єдиний безперервний полінуклеотидний ланцюг. В еукаріот генетичний матеріал розподілений по хромосомах й у кожній хромосомі теж утворює одну довгий полінуклеотидний ланцюг.
Полінуклеотидні нитки ДНК, що містяться в хромосомах еукаріот, у геномі бактерій і вірусів або плазмід (у деяких вірусів - РНК), підрозділяються на функціональні відрізки, називані генами (спадкоємні задатки).
Розрізняють:
1) структурні гени, у яких закодована інформація для синтезу ферментних і структурних білків;
2) гени з інформацією для синтезу t PHK;
3) гени з інформацією для синтезу r РНК;
4) специфічні регуляторні ділянки, такі як промотори й оператори;
5) поділяючі ділянки між генами (спейсори);
6) ділянки з невідомою функцією.
Визначено послідовності для деяких повних геномів і для багатьох частин геномів і зроблені висновки про їхні функції. Гени відділені один від одного спейсорами. Крім того, є ділянки, "розпізнавані" певними молекулами, такими як промотор й оператор, для регуляції активності генів. В еукаріотів крім структурних генів, які розташовані в хромосомах лінійно, існують ділянки з повторюваними послідовностями.
5.1 Дія генів
Першим кроком на шляху до формування ознаки є транскрипція - "переписування" нуклеотидних послідовностей певних ділянок ДНК у формі m PHK. Після цього РНК-транскрипт (пре-РНК) може модифікуватися в результаті посттранскрипційних процесів.
На рибосомах здійснюється трансляція - переклад нуклеотидної послідовності m PHK в амінокислотну послідовність поліпептиду. Поряд з багатьма білками для цього процесу необхідні також t PHK й r РНК.
Ті або інші ознаки клітини або організму з'являються завдяки тому, що утворюються специфічні структурні білки або ж ферменти, відповідальні за певні етапи процесів синтезу або розпаду. У результаті з попередників через проміжні етапи утворюються кінцеві продукти, від яких і залежить прояв специфічних ознак, тобто функціональних здатностей (рис. 5.1).
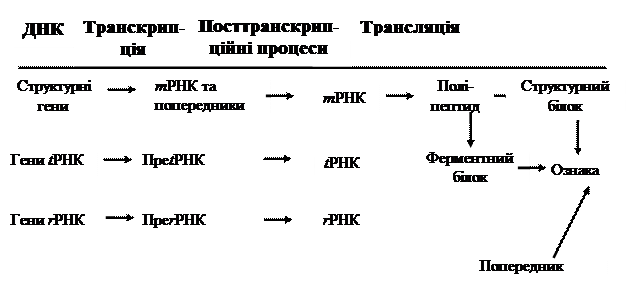
Рис. 5.1. Реалізація генетичної інформації (схема)
5.1.1 Транскрипція
При транскрипції рибонуклеозиди - цитидин, гуанозин, уридин й аденозин - С, G, U й А, синтезовані в процесі клітинного метаболізму у формі рибонуклеозидтрифосфатів (rNTP) СТР, GTР, UТР й АТР, пристроюються до компліментарних основ ДНК, а саме C до G, G до C, U до А и А до Т. Транскрипція йде від початку транскрипційної одиниці до її кінця. ДНК-матрицею служить так називаний кодогенний ланцюг ДНК, на якому транскрипційні одиниці транскрибуються в напрямку 3'®5'. Рибонуклеозидтрифосфати (rNTP) за допомогою ферментів - ДНК--залежних РНК-полімераз - зв'язуються (з наступним відщіпленням пірофосфату) у напрямку 5'® 3' у ланцюг РНК (рис. 5.2, А). Цей фермент повинен не тільки забезпечувати правильне утворення пар і зв'язування rNTP у ланцюг, але й знаходити належне місце початку транскрипції, вибирати кодогенний ланцюг і розділяти два ланцюги ДНК біля тих пар основ, де відбувається транскрипція. Синтез РНК закінчується на термінаторній ділянці ДНК. Транскрипція активного гена здійснюється так, що на транскрипційній одиниці одночасно перебуває багато молекул полімерази зі зростаючими ланцюгами РНК (рис. 5.2, Б). В еукаріот для транскрипції ядерної ДНК існують три різні РНК-полімерази; структурні гени транскрибуються полімеразою ІІ.
В одній певній фазі життєвого циклу транскрипції піддається лише близько 10% структурних генів; інші гени неактивні, але можуть стати активними в інших фазах.
В еукаріот транскрипція відбувається в клітинному ядрі, а трансляція - у цитоплазмі на рибосомах. Пре- m PHK у формі рибонуклеопротеїдних часток попадає в цитоплазму й перетерплює ряд змін, які називають посттранскрипційними процесами. Як у прокаріот, так й в еукаріот первинний транскрипт (пре- m PHK) у більшості випадків довше, ніж послідовність нуклеотидів, що відповідає кінцевому продукту (поліпептиду, t PHK, r РНК). Готова m PHK починається із вступної послідовності (лідера), потім йде ділянка, що несе інформацію для генного продукту, і кінцева послідовність (трейлер).
| А | 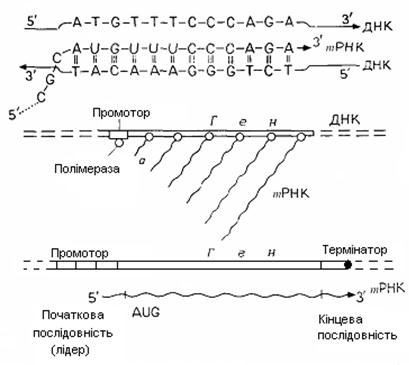
|
| Б | |
| В |
Рис. 5.2. Транскрипція. А. Синтез m PHK у початковій частині гена.
Б. Повторний старт синтезу m PHK на гені.
В. Транскрибуєма ділянка генетичного матеріалу
В еукаріот часто зустрічаються різноманітні розходження між первинним транскриптом і РНК, що надходить для трансляції. Після транскрипції можуть відбуватися наступні зміни (у сукупності називані також процесінгом:
а) утворення "ковпачка" (метилювання й формування спеціальної структури на 5'-кінці);
б) метилювання внутрішніх основ;
в) поділ всієї послідовності на частини або відщіплення певних частин;
г) видалення внутрішніх ділянок і з'єднання послідовностей, що залишилися, - сплайсінг;
д) додавання poly (А) до 3'-кінцю.
Утворення ковпачка й внутрішнє метилювання пре- m РНК типово для еукаріот. В еукаріот у послідовності нуклеотидів ДНК з інформацією для послідовності амінокислот деяких поліпептидів включені відрізки, що не містять інформації, так називані інтрони. Спочатку утворюється транскрипт всієї послідовності (пре- m PHK), а потім відбувається сплайсінг й інформативні відрізки поєднуються в одну безперервну послідовність - m РНК. Припускають, що частина посттранскрипційних процесів має значення для регулювання дії генів. Для початку транскрипції велике значення має так званий промотор. Це передуюча гену послідовність, що складається приблизно з 80 нуклеотидів, яку розпізнає й з якої зв'язується фермент РНК-полімераза.
5.1.2 Трансляція
При трансляції нуклеотидна послідовність t РНК переводиться в амінокислотну послідовність поліпептидного ланцюга. Ключем для перекладу послідовності нуклеотидів у послідовність амінокислот служить генетичний код. Як при перекладі з однієї мови на іншу необхідно знати слова, так і тут варто знати, як кожна амінокислота закодована в нуклеотидах.
В t РНК є 4 різних нуклеотида: С, G, U й А. У біосинтезі білка беруть участь 20 амінокислот. Щоб їх закодувати, необхідно щонайменше 20 різних знаків. При чотирьох різних нуклеотидах це можливо у випадку складання знаків із трьох нуклеотидів; таких знаків може бути 43 = 64. Три нуклеотида, що утворять кодовий знак, називають триплетом.
Генетичний код удалося з'ясувати завдяки роботам дослідницьких груп Ніренберга й Очоа (табл. 5.1). Результати цих робіт стали одним із самих значних кроків у розумінні життєвих процесів.
Ці результати можна коротко резюмувати в такий спосіб:
1. Генетичний код являє собою триплетний код. Триплет m РНК одержав назву кодона.
2. Генетичний код є виродженим кодом, тобто одній амінокислоті, як правило, відповідає більш ніж один кодон. У кодонах для однієї амінокислоти перші два нуклеотида найчастіше однакові, а третій варіює.
3. Нуклеотидна послідовність зчитується в одному напрямку підряд, триплет за триплетом. Кодони не перекриваються.
4. AUG являє собою стартовий кодон.
5. UAG (amber), UAA (ochre) і UGA (opal) - кодони-термінатори.
6. Генетичний код універсальний, він єдиний для всіх організмів і вірусів.
Таблиця 5.1
Генетичний код (кодони t РНК для амінокислот)
| Амінокислота | Кодони для амінокислоти |
| Фенілаланін | 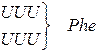
|
| Лейцин | 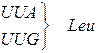
|
| Аланін | 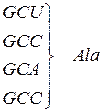
|
| Валін | 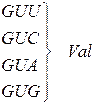
|
| Серин | 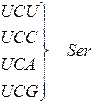
|
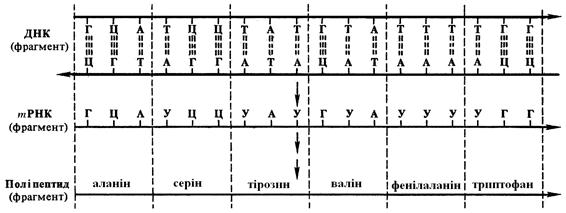
Рис. 5.3. Генетичний код. Співвідношення послідовності триплетів ДНК, РНК і мономерів (амінокислот) у білковій молекулі
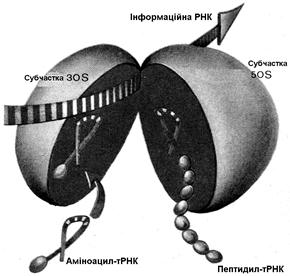
Трансляція здійснюється на рибосомі. t РНК прикріплюється до малої субчастки, зв'язування амінокислот відбувається на великий субчастці. Крім інформації для амінокислотної послідовності, t РНК містить початкову (лідер) і кінцеву (трейлер) послідовності. Частина початкової послідовності необхідна для зв'язування з малою субчасткою рибосоми. "Ковпачку" в еукаріот приписують роль у впізнаванні рибосоми й у зв'язуванні з малої субчасткою.
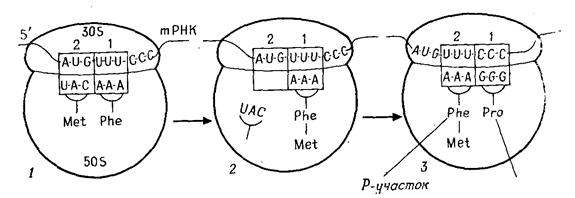
Трансляція починається зі стартового кодона AUG (рис. 5.4). На малій субчастці є особлива ділянка (позиція 1), де триплет підготовлюється до трансляції. Тому що генетичний код не має "ком", початок зчитування повинен визначатися точно, і для цієї мети служить стартовий кодон AUG. Потім на те ж місце для зчитування безупинно один за одним встають наступні триплети.
Роль посередника між кодоном m РНК й амінокислотою грає t PHK. Для кожної амінокислоти існує, щонайменше, одна t PHK. Вона виконує в основному два завдання: вибирає з 20 амінокислот специфічну для неї й знаходить за допомогою свого антикодону (рис. 2.8. й 5.4) відповідний кодон m РНК за принципом спарювання основ. Та t PHK, що, підійшовши до малої субчастки, утворить зв'язок кодон-антикодон, одночасно передає свою амінокислоту на аміноацильну ділянку великої субчастки. До кодона AUG "підходить" антикодон тільки тієї t PHK, що переносить метіонін (Met-t PHK). Тому насамперед до рибосоми доставляється метіонін. Потім одночасно кодон AUG переходить із позиції 1 у позицію 2 на малій субчастці, а Met - t PHK - на пептидильну ділянку великої субчастки. На малій субчастці в позиції 1 стоїть тепер наступний кодон (на рис. 5.4 - UUU), готовий до зв'язування з антикодоном. До кодона UUU "підходить" антикодон Phe-t PHK, і на аминоацильну ділянку великої субчастки попадає амінокислота фенілаланін. Карбоксильна група метіоніну, що раніше була пов'язана з t PHK, приєднується до аміногрупи фенілаланіну, і утворюється дипептид, Met-t PHK звільняється й готова тепер зв'язати нову молекулу метіоніну. Коли другий кодон, UUU, переходить у позицію 2, позиція 1 звільняється для наступного кодона (на рис. 5.4. - ССС). Знову відбувається спарювання основ кодона й антикодон. На аміноацильну ділянку великої субчастки попадає амінокислота пролін. Утворюється пептидний зв'язок між фенілаланіном і проліном. У такий спосіб зчитується триплет за триплетом. Послідовність кодонів в t РНК визначає послідовність амінокислот у поліпептиді. Утворення пептидних зв'язків припиняється, коли на ділянці трансляції з'являється кодон-термінатор t РНК. Для нього не існує t PHK, і на аміноацильну ділянку не попадає ніяка амінокислота.
А
Б
Рис. 5.4. Трансляція. А. Об’єднання субчасток рибосоми на m РНК (схема).
Б. Утворення білків на рибосомі
Обидві субчастки поєднуються в здатну функціонувати рибосому тільки тоді, коли до малої субчастки вже прикріпилася t РНК, а до останньої приєдналася Met-t PHK. Цю початкову фазу трансляції називають ініціацією. Зв'язування наступних амінокислот у поліпептид відбувається у фазі елонгації (подовження ланцюга). У фазі термінації синтез поліпептидного ланцюга закінчується при участі звільняючих ланцюг факторів. t РНК одночасно зчитується декількома рибосомами ("полісоми"). Ще до того, як вона буде повністю трансльована, її початок попадає на наступну рибосому й там теж транслюється.
На рибосомі правильне розміщення амінокислот у поліпептидному ланцюзі здійснюється завдяки спарюванню основ між антикодоном t PHK і кодоном m РНК. За зв'язування амінокислоти з відповідною t PHK відповідає специфічний фермент - синтетаза. Кожна амінокислота має свою специфічну синтетазу (або декілька таких синтетаз). Всі t PHK мають акцепторний кінець, до якого приєднується амінокислота (див. рис. 2.8). Приєднання амінокислот до специфічних t PHK - необхідна передумова безпомилкового ходу біосинтезу білків, тому що кожна амінокислота, пов'язана з t PHK, включається в пептидний ланцюг відповідно до антикодону цієї t PHK.
5.2 Від поліпептиду до ознаки
Поліпептидні ланцюги, що утворюються при біосинтезі білка, визначають ознаки клітини й організму, формуючи білкові структури або управляючи процесами метаболізму як ферменти. Як структуроутворюючі елементи поліпептиди беруть участь у побудові мембран, хроматину, рибосом, мікрофіламентів і мікротрубочок. Як ферментні білки вони управляють всіма важливими процесами, які забезпечують формування клітин й організмів, підтримку їх структур і функцій, розвиток, клітинний поділ і розмноження. Переважна більшість метаболічних реакцій перебуває під контролем ферментів й, отже, генів. Відповідне відношення між геном і ферментом встановлене в 1944 р. Бідлом і Татумом і ввійшло в біологію як гіпотеза "один ген - один фермент". Всі життєві процеси, а виходить, і ознаки організму (колір волосся, забарвлення квітів, ріст, продуктивність і всі інші функції) визначаються генами.
Сукупність всіх ознак організму називається фенотипом. Фенотип залежить від:
1) генотипу, тобто від наявних генів і взаємодії алелей;
2) внутрішнього й зовнішнього середовища, що впливає на активність генів й обумовлює регуляторні процеси й модифікації.
5.3 Регуляція генної активності
Якісний і кількісний склад білків в одній клітині (у різний час) і в різних клітинах організму може бути досить різним. Розходження спричиняються процесами, що відбуваються при синтезі білків de novo - на стадіях транскрипції, посттранскрипції або трансляції, і впливами на вже наявні білки або їх попередники.
5.3.1 Регулювання транскрипції
Інтенсивність транскрипції безпосередньо визначається промотором. Конститутивні гени активні завжди, і їхні продукти увесь час утворюються в однакових кількостях; активність цих генів залежить від взаємодії РНК-полімерази із промотором. Крім того, на транскрипцію, обумовлену промотором, можуть впливати деякі побічні фактори. Дія регульованих генів може змінюватися залежно від зовнішнього середовища й стадії розвитку в результаті регуляторних процесів. Наявна ДНК обумовлює тільки потенційні можливості, реалізація ж їх залежить від внутрішнього й зовнішнього середовища.
Промотор розпізнається ферментом і вказує місце, де повинна починатися транскрипція. За цю функцію відповідальні дві послідовності, що лежать на відстані 35 й 10 нуклеотидів від місця старту транскрипції. Позитивною регуляцією транскрипції називають механізм, при якому регулюючі фактори активують промотор. Про негативну регуляцію транскрипції говорять у тих випадках, коли регулюючі фактори запобігають транскрипції.
У бактерій 5-10% всіх генів утворюють функціональні групи. Перед групою структурних генів, продукти яких належать переважно до одного метаболічного шляху, перебувають загальний для них промотор і загальний оператор. Таку функціональну одиницю називають опероном (рис. 5.5). На структурних генах, що належать до одного оперона, утворюється одна загальна молекула t РНК (поліцистронна t РНК), так що всі ці гени одночасно активні або неактивні.
Інтенсивність транскрипції регулюється в результаті взаємодії оператора з білком-репресором. Оператор - це послідовність приблизно з 30 нуклеотидів, яку "розпізнає" репресор.
Ця послідовність має дзеркальну симетрію (рис. 5.4, В). Структуру репресора кодує ген-регулятор. Репресор являє собою алостеричний білок, що може приймати дві різні конформації. Молекула репресора має дві специфічних ділянки: одну для приєднання до оператора й одну для зв'язування ефектора. Приєднуючись до оператора, репресор блокує транскрипцію. Він викликає негативний ефект, тому такий тип регуляції називають негативним контролем. Зв'язування ефектора зі специфічною ділянкою змінює просторову структуру репресора таким чином, що останній при синтезі репресибельного ферменту
| А | 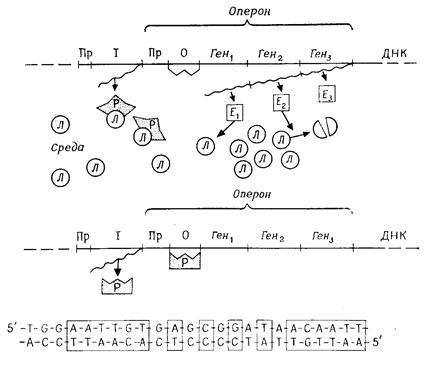
|
| Б | |
| В |
Рис. 5.5. Регуляція транскрипції. Приклад: область лактозного оперона в Escherіchіa colі. А. У присутності лактози. Б. Без лактози. Пр – промотор; О –оператор; І – регуляторний ген; Ген1 Ген2, Ген3 –структурні гени; Р – репресор; Л – молекула лактози; хвиляста лінія – t РНК. В. Послідовність lас -оператора. У рамках: комплементарно-симетричні ділянки
блокує оператор, а при синтезі індуцибельного ферменту звільняє оператор.
Регуляція за допомогою системи репресор - оператор відома для деяких катаболічних й анаболічних процесів у бактерій. Регуляція на рівні оператора називається моделлю Жакоба-Моно по імені вчених, які її відкрили.
У випадку катаболічних процесів - при розщепленні субстратів - така регуляція зручна тому, що фермент, що розщеплює, утворюється тільки тоді, коли є субстрат; наявність субстрату індукує синтез ферменту, і такі ферменти називають індуцибельними. Субстрат (або один із продуктів його обміну) служить ефектором репресора.
Особливо добре вивчена регуляція розщеплення лактози в Escherіchіa colі (рис. 5.4). Якщо лактози в середовищі немає, то репресор блокує оператор і ферменти лактозного оперона (lac -оперона) не синтезуються. Якщо ж лактоза в середовищі є, репресор взаємодіє з нею й змінює при цьому свою конформацію так, що більше не може блокувати оператор. У результаті на lac-опероні утворюється m РНК, і завдяки цьому синтезуються ферменти, що розщеплюють лактозу.
У випадку анаболічних процесів - при синтезі компонентів клітини - за допомогою регуляції досягається те, що вихідні речовини не накопичуються в надлишковій кількості. Наприклад, синтез тієї або іншої амінокислоти, коли в клітині її вже досить, "вимикаються" (відповідні ферменти називають репресибельними).
Регуляція генної активності в еукаріотів набагато складніше. Розходження між про- і еукаріотами пов'язане з тим, що в еукаріот поряд з регуляторними процесами, що впливають (як й у прокаріот) на функції й життєвий цикл окремої клітини, існують і такі процеси, які впливають на розвиток усього організму. Крім того, у прокаріот транскрипція й трансляція звичайно відбуваються безпосередньо одна за одною, а в еукаріот транскрипт повинен перейти з ядра в цитоплазму на рибосоми. Очевидно, поряд з регулюванням транскрипції в еукаріот значну роль грає регуляція посттранскрипційних процесів. У ссавців важливе значення має дія гормонів. За допомогою методів молекулярної біології була досліджена регуляторна дія гістонів і негістонових хромосомних білків. Для активації транскрипції велике значення має зміна конформації хроматину. Крім того, надзвичайно важлива роль інтронів у регуляторних механізмах.
5.3.2 Регулювання трансляції
У прокаріот m РНК містить перед стартовим кодоном так звану послідовність Шайна-Дальгарно з 6-8 нуклеотидів (найчастіше AGGAGGU); ця область бере участь у зв'язуванні m РНК із субчасткою рибосоми. Показано, що ефективність трансляції залежить від лідерної послідовності й, імовірно, визначається вторинною структурою цієї ділянки.
В еукаріот регулювання трансляції відбувається на етапі ініціації. На відміну від прокаріот, що мають тільки три фактори ініціації, в еукаріот відомо 7-8 таких факторів. Вони побудовані більш складно й мають істотно більшу молекулярну масу.
5.4 Модифікації
Модифікаціями називають неспадкові зміни фенотипу, викликані впливом навколишнього середовища. Генотип залишається незміненим.
Формування ознаки - ланцюг процесів, що йде від гена через m PHK, поліпептид і фермент, - протікає нормально тільки в тому випадку, якщо в розпорядженні клітини є всі необхідні вихідні речовини, належне джерело енергії й підходящі умови для реакцій (такі як рН, концентрації іонів і т.п.). Таким чином, середовище повинно забезпечити умови, необхідні для формування ознаки.
Картопля, поміщена в підвал, не утворить зелених пластид, хоча гени для цього є. На світлі паростки, утворені такою же картоплею, зеленіють. Синтез хлорофілу залежить, таким чином, не тільки від відповідних генів, але й від зовнішнього фактора - світла.
Особливо серйозно позначаються ті фактори зовнішнього середовища, від яких залежить харчування. Погано вдобрені поля не приносять великого врожаю; тварини, що не одержували досить їжі, не мають достатню продуктивність або працездатність.
Гени визначають норму реакції, а від зовнішнього середовища залежить, який варіант у межах цієї норми реакції реалізується в цьому випадку (рис. 5.6).
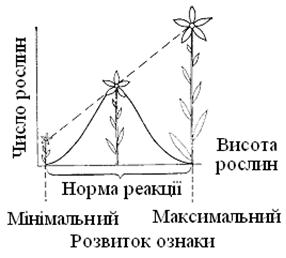
Рис. 5.6. Генетично детермінована норма реакції певного виду рослин
У результаті селекції сорти сільськогосподарських рослин мають потенційну здатність давати більші врожаї. Однак ця здатність, обумовлена генетично, може бути реалізована повністю тільки тоді, коли агротехніка (обробка й удобрення ґрунту, забезпечення водою) створює належні передумови для цього.
На рис. 5.6 крива модифікацій відбиває частоту зустрічальності різних форм. На організм рідко впливає відразу багато дуже несприятливих або дуже сприятливих факторів середовища; тому індивідуумів з мінімальним або максимальним розвитком якої-небудь ознаки буває мало, а частіше всього зустрічаються проміжні форми.
Точно так само й у людей індивідуальність повністю проявляється лише в результаті взаємодії генетичних задатків і зовнішнього середовища, причому до діючих факторів середовища поряд з харчуванням і фізичними умовами ставляться й грають настільки ж вирішальну роль виховання й суспільні відносини.
5.5 Взаємини алелей
Ген - це послідовність нуклеотидів з певною біологічною активністю. Кожен ген може бути представлений різними формами, які звуться алелі. У гаплоїдних організмів кожен ген представлений тільки одним алелем, і на основі укладеної в ньому інформації формується відповідна ознака.
У диплоїдних клітин кожен ген представлений двічі. Якщо це в обох випадках той самий алель, клітину називають гомозиготною, а якщо це різні алелі - гетерозиготною.
У гетерозиготних організмів алелі одного гена можуть взаємодіяти різним чином: найчастіше алель дикого типу (нормальний) буває домінантним, а мутантний алель - рецесивним (тобто подавленим). Між домінантністю й рецесивністю існують всі переходи. Зокрема, нерідкий прояв ознаки в гетерозиготі буває проміжним стосовно крайніх форм, що реалізується в обох гомозиготах. Наприклад, у гетерозиготи може виявлятися тільки 50% ферментативної активності, властивої гомозиготі.
5.6 Полігенне спадкування й плейотропія
Коли у визначенні однієї ознаки беруть участь кілька генів, це явище називають полігенією. Наприклад, ряд амінокислот утворюється в результаті декількох реакцій при участі декількох генів. Жоден з генів, що контролюють різні етапи синтезу, сам по собі не може забезпечити прояв даної ознаки - для здійснення синтезу необхідна взаємодія всіх цих генів (компліментарна полігенія).
Кількісні ознаки, такі як розміри тіла, вміст певних речовин або врожайність, часто залежать від багатьох генів таким чином, що кожен окремий ген має лише слабку дію й для повного вираження ознаки необхідна велика кількість генів з подібною дією (адитивна полігенія).
Залежність розвитку декількох ознак від одного гена називається плейотропією. Плейотропна дія гена проявляється й тоді, коли який-небудь продукт діє в різних частинах організму й може обумовлювати різні ознаки. Наприклад, у рослин фіолетово-червоне забарвлення квітів, плодів, листів і проростків залежить від присутності одного і того ж пігменту - антоціану, а тому й від одного гена.
Запитання для самоперевірки
1. Які види генів відомі?
2. В яких процесах відбувається формування ознак організму?
3. Які ферменти здійснюють транскрипцію?
4. Що таке пост транскрипційні процеси?
5. Які властивості генетичного коду?
6. Що таке кодон і антикодон?
7. Де відбувається синтез білків у рослинних та тваринних клітинах?
8. Як відбувається трансляція?
9. Як відбувається негативна регуляція генної активності (схема Жакоба-Моно)?
10. Які взаємини алелей відомі?
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 8587; Нарушение авторских прав?; Мы поможем в написании вашей работы!