КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Profesor universitar D.Nedeoglo
|
|
|
|
Л е к ц і я 2. Основні поняття в екології. Екологічна система, біогеоценоз та його структура. Трофічні ланцюги живлення. Екологічні фактори. Лімітуючий фактор. Біотичні та абіотичні фактори.
Plan
1. The syllable as an integral part of the word. Types of syllables in English.
2. The syllable formation theories.
3. Basic rules of syllabification in English.
4. Division into syllables in writing.
1. The syllableis a group of sounds that are pronounced together. The syllable is one or more speech sounds forming a single uninterrupted of utterance unit which may be a commonly recognized subdivision of a word.
The problem of the syllabic structure of words has two aspects:
1) syllable formation(складоутворення)
2) syllable division/separation (складоподіл).
Articulatorily,the syllable is the minimal articulatory unit of the utterance. Auditorily,the syllable is the smallest unit of perception: the listener identifies the whole of the syllable and after that the sounds which it contains. Phonologicallyit is a structural unit which consists of a sequence of one or some phonemes of a language in numbers and arrangements permitted by the given language. Phonologically it performs three functions:
1. The constitutive function(конститутивна функція): syllables constitute words through the combination of their stress-loudness, duration-length, pitch-tone.
2. The distinctive function(смислорозрізнювальна / дистинктивна функція): the difference in the place of a syllabic boundary differentiates the meanings of the words and phrases:
e.g. a 'name – an 'aim; kids 'kin - kid 'skin; my 'train - might 'rain
3. The identificatory function (ідентифікативна функція): the listener can understand the exact meaning of the utterance only when the correct syllabic boundary is perceived: e.g. peace talks - pea stalks.
In English the syllable is formed:
1) by any vowel alone or in combination with one or more consonants - not more than 3 preceding and not more than 4 following it, e.g. are [a:], we [wi:], it [it], sixths [siksӨs].
2) by a word final sonorants [n], [l], [m] immediately preceded by a consonant: e.g. rhythm ['rið. m], garden I'ga:. dn].
According to the placement of vowels and consonants the following types of syllables are distinguished:
| 1. Fully open | V ore, or |
| 2. Fully closed (V between C) | CVC fat CCVC place CVCC fact CCCVCC street CVCCC facts CVCCCC sixths |
| 3. Covered at the beginning (one C or a sequence of C precede a vowel) | CV too CCV spy CCCV straw |
| 4. Covered at the end (one C or more complete the syllable) | VC on VCC act VCCC acts |
Structurally, the commonest types of the syllable in English are VC; CVC.
Syllables can be also designated:
1) by the position in the word:
from the beginning - INITIAL (початковий), MEDIAL (серединний), FINAL (фінальний/кінцевий) or
from the end - ULTIMATE (останній), PENULTIMATE (передостанній/другий від кінця), ANTEPENULTIMATE (третій від кінця);
2) by the position in relation to stress:
PRETONIC (переднаголошений), TONIC (наголошений), POSTTONIC (післянаголошений)
Any syllable which is not tonic is ATONIC (ненаголошений).
e.g. tre - men - dous
initial medial final
antepenultimate penultimate ultimate
pretonic tonic posttonic
2. There have been suggested several theories to explain the mechanism of syllable formation and syllable division in different languages.
The first theory in chronological order is the chest pulse/breath puff (теорія видиху). According to this theory, syllables are formed by breath-puffs. At present this theory is considered as not scientifically grounded; it has been proved experimentally that speakers take a new breath at the end of a syntagm and no increase of breath force necessarily accompanies every syllable.
The second in chronological order comes the relative sonority theory/the prominence theory (теорія відносної сонорності). It is based upon the fact that each sound has a different carrying power which is based on their sonority. The sonority of a sound is its relative loudness compared to other sounds, everything else (pitch, etc.) being equal.
The creator of this theory, the Danish linguist Otto Jespersen, has proved that the least sonorous sounds which have the least carrying power, are those for which the mouth is closed (voiceless oral stops), while the most sonorous sounds are those for which the mouth is wide open (low vowels). All other sounds are ranked in between these two extreme points of the sonority scale: (from the highest degree to the lowest):
1. Low vowels (a:, ……).
2. High vowels (i:, i ……)
3. Semivowels (j, w)
4. Liquids (l, r)
5. Nasals (m, n, ŋ)
6. Fricatives (voiced) (v, z, ð)
7. Fricatives (voiceless) (f, Ө, s)
8. Oral stops (voiced) (b, d, g)
9. Oral stops (voiceless) (p, t, k).
By this theory the syllable is treated as the combination of a more sonorous sound with a less sonorous one. All the sounds with the greatest degree of sonority (vowels and sonorants) are at the peak of the syllable, by which the syllable may be marked as a unit, because the rest of the sounds surrounding the peak cling to it.
The sonority theory helps establish the number of syllables in a word, but it's difficult to find the syllable boundary using this syllable as it doesn't explain the mechanism of syllable division.
The muscular tension theory (теорія м’язового напруження) was put forward by the French linguist Michaelle Grammont and supported and further developed by the Russian linguist Lev V. Scherba.
Academician Lev Volodymyrovych Scherba explained syllable formation by muscular tension impulses and three types of consonants. In speaking, muscular tension impulses follow one another. Each impulse has its strongest point - the peak of prominence, - and its weakest prominence - the valley of prominence. Valleys of prominence correspond to points of syllabic division. The end of one syllable and the beginning of the next one can be ascertained by determining the type of consonants which take part in forming the syllables.
Consonants may be pronounced:
(1) initially strong - the beginning of a consonant may be more energetic, while the end may be weaker;
(2) finally strong - the beginning of the consonant may be weak, and its end – more energetic;
(3) geminate or double - both the beginning and the end are energetic with a weakening of muscular tension in the middle, acoustically, they give the impression of two consonants.
The more energetic part of a consonant is attached to a vowel, so that initially strong C occurs at the end of a close syllable, while finally strong C occurs at the beginning of a syllable. This theory again does not give a complete explanation of the syllable division mechanism.
The problem of syllable formation and syllable division in English is open for further investigation.
3. The division of a word into syllables is called syllabification. The question of syllabification in English is controversial: different phoneticians hold different views about it. It is generally agreed that phonetic syllable divisions must be such as to avoid (as far as possible) creating consonant clusters which are not found in words in isolation. Thus it may be argued that candy should be ['kæn. di] or ['kænd. i] but not ['kæ. ndi] since [nd] is not a possible initial consonant cluster in English. This principle is called the phonotactic constraint(фонотактичне обмеження) on syllabification.
The following rules of phonetic(spoken) syllable divisionare adopted in LPD-2000:
1. A syllable boundaryis found wherever there is a word boundary,and also coincides with the morphological boundary between elements in a compound:
displace [,dls 'pleis], become [bi 'k m], countless ['kaunt ləs], hardware ['ha:d weə]
2. Consonantsare syllabified with whichever of the two adjacent vowels is more strongly stressed, e.g. farmer ['fa:m ə], agenda [ə 'd3ənd ə].
If they are both unstressed, it goes with the leftward one: e.g. cinema ['sin əm ə], delicious [di 'liS əs], deliberate [di 'lib ər ət].
3. The English diphthongsare unisyllabic, they make one vowel phoneme, while the so called triphthongs are disyllabic, because they consist of a diphthong + the neutral vowel/schwa:
table science flower
CV-CS CV-VSC CSV-V
4. The English affricates [t ], [d3] cannot be split: catching ['kæt. iŋ].
4. Phonetic (spoken) syllables must not be confused with orthographic (written) syllables. An orthgraphic syllable is a group of letters in spelling. Syllables in writing are also called syllabographs.
When a word is split across two lines of writing, it should be broken at an orthographic syllable boundary. Parts of phonetic and orthographic syllables do not always coincide: worker ['w3:k.ə] CVC - V = two phonetic syllables and one syllabograph.
A most general rule claims that division of words into syllables in writing is based on the morphological principlewhich demands that the part of a word which is separated should be either a prefix, or a suffix or a root (morphograph), e.g. pic - ture ['pik tS ə].
Compound wordscan be divided according to their meaning: hot -dog; spot-light.
It is not possible to divide a word within a phonetic syllable.
A suffix of two syllables such as -ABLE, -ABLY, -FULLY cannot be divided in writing, e.g. reli-able, lov-ably, beauti-fully. If there are two or three consonants before - ING, these consonants may be separated in writing: gras-ping, puz-zling.
With the exception of -LY, a word cannot be divided so that an ending of two letters such as -ED, -ER, -IC begins the next line, e.g. worked, teacher, hectic, BUT: cold-ly, bold-ly.
A word of one phonetic syllable, a word of less than five letters cannot be divided into syllabographgs, e.g. piece [pi:s], time [taim].
Природне середовище – це все живе й неживе, що оточує організми і з чим вони взаємодіють. Розрізняють повітряне, водне та ґрунтове середовище, таким середовищем може бути і тіло іншого організму (для паразитуючих організмів).
Вид (біологічний) – сукупність організмів із спорідненими морфологічними ознаками, які можуть схрещуватись один з одним та мають спільний генофонд. Це основна структурна одиниця в системі живих організмів (в біології).
Популяція – сукупність особин одного виду з однаковим генофондом, яка живе на спільній території протягом довгого часу.
Стабільний стан в екології називають гомеостазом. Гомеостаз – це стан внутрішньої динамічної рівноваги природної системи, що підтримується регулярним відновленням її основних елементів і речовинно-енергетичного складу, а також постійним функціональним саморегулюванням компонентів.
Основною елементарною (функціональною) одиницею екології є екосистема.
Екосистемою називають будь-яку сукупність організмів і неорганічних компонентів, в якій може здійснюватись кругообіг речовин. Термін „екосистема” був запропонований у 1935 р. англійським екологом А. Тенслі.
Розрізняють мікроекосистеми (пеньок з грибами, болітце), мезоекосистеми (ділянка лісу, озеро), та макроекосистеми (континент, океан). Глобальною екосистемою є біосфера нашої планети.
Угруповання живих організмів утворює з неорганічним середовищем певну систему, в якій потік атомів, викликаний життєдіяльністю організмів, має тенденцію замикатися як кругообіг (закон біогенної міграції атомів – Вернадського). Для підтримання круговороту речовин в системі необхідна наявність запасу неорганічних молекул в освоюваній формі і трьох функціонально різних екологічних груп організмів: продуцентів, консументів і редуцентів:
Продуцентами виступають автотрофні організми, які здатні будувати своє тіло за рахунок неорганічних сполук (в основному, рослини).
Консументи – це гетеротрофні організми, які вживають органічну речовину продуцентів або інших консументів і трансформують її у нові форми (більшість тварин).
Редуценти живуть за рахунок мертвої органічної речовини, переводячи її знову в неорганічні сполуки (переважно, мікроорганізми та гриби).
Для екологічної системи характерні наступні ознаки:
1) екосистема обов'язково складається з живої та неживої складових;
2) у рамках екосистеми відбувається повний цикл, починаючи з утворення органічної речовини та закінчуючи його розкладанням на неорганічні складові;
3) екосистема зберігає стійкість протягом певного часу, що забезпечується певною структурою біотичного та абіотичного компонентів;
4) більш прості екосистеми входять у більш складно організовані. При цьому реалізується ієрархія їх організації. Важливим наслідком ієрархічної організації екосистем є те, що у ході організації окремих компонентів екосистеми у більш складні блоки, які, в свою чергу, об'єднуються в системи, у цих нових функціональних одиниць утворюються нові властивості, яких не було на попередніх рівнях. Така наявність у системного цілого особливих ознак, яких немає у підсистемних блоків, називають емерджентністю.
Коротко закон емерджентності можна сформулювати так: ціле більше, ніж сума його частин. Тому емерджентність екосистем означає не просто перехід кількості в якість, а являє собою особливий тип інтеграції. Емерджентні властивості певного екологічного рівня не можна передбачити, знаючи лише ознаки та властивості компонентів, які складають цей рівень – його потрібно вивчити безпосередньо. Наприклад, молекула має інші властивості, ніж атоми, що її складають. Тому, вивчаючи скупчення атомів, ми не зможемо передбачити властивості молекул, що можуть з них утворитися. Так само, скупчення окремих молекул (або навіть органів) не дасть властивостей цілого організму і т.д.
Принцип емерджентності має важливе значення для екологічного мислення: так, єдине дерево не може скласти лісу, декілька дерев – також. Ліс виникає тільки при певних умовах – достатній густині деревостою, певній флорі та фауні, сформованих співтовариствах певних організмів, які живуть на його території та багатьох інших умовах. Таким чином, емерджентні властивості виникають в результаті зміни природи компонентів, а не в результаті зміни кількості компонентів. Частини не злипаються, а інтегруються, забезпечуючи виникнення нових, відсутніх до цього властивостей.
Таким чином, структуру природи слід сприймати як єдине ціле, що складається з вкладених одна в одну екосистем, найбільшою з яких є глобальна екосистема – біосфера. У її межах відбувається постійний обмін речовиною та енергією між всіма живими та неживими компонентами в планетарних масштабах.
Паралельно розвиткові концепції екосистем успішно розвивається вчення про біогеоценози, автором якого був академік В.М. Сукачов (1942). "Екосистема" та "біогеоценоз" – близькі за своєю суттю поняття, однак якщо перше з них придатне для позначення систем, які забезпечують кругообіг будь-якого рангу, то " біогеоценоз" – поняття територіальне і вживається для позначення ділянок суші, зайнятих певними одиницями рослинного покриву – фітоценозами. Наука біогеоценологія виросла з геоботаніки і спрямована на вивчення функціонування екосистем у конкретних умовах ландшафту, залежно від властивостей ґрунту, рельєфу, а також характеру оточення біогеоценозу і, зокрема, його первинних компонентів – гірської породи, тварин, рослин, мікроорганізмів.
Біогеоценологія розглядає поверхню Землі як мережу біогеоценозів-сосудів, пов'язаних між собою через міграцію речовин, але хоч і не однаковою мірою, та все ж автономних і специфічних за своїми кругообігами. Конкретні умови ділянки, зайнятої біогеоценозом, надають їй своєрідності, вирізняючи з-поміж інших, схожих за типом.
Компоненти біогеоценозу – біотоп та біоценоз. Біотоп – однорідний за біотичними факторами середовища простір, зайнятий біоценозом, а біоценоз – спільнота організмів, що мешкають у межах одного біотопу. Поняття „біоценоз” є умовним, так як поза середовищем існування організми жити не можуть, але ним зручно користуватися в процесі вивчення екологічних зв'язків між організмами.
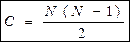
Трофічні ланцюги живлення
Ряди, в яких можна простежити шляхи витрачання початкової дози енергії, називають ланцюгами живлення – система передачі та перерозподілу речовини та (частково) енергії, яка реалізується шляхом поїдання одних видів організмів іншими.
Місце кожної ланки в ланцюгу живлення називають трофічними рівнем:
I трофічний рівень – це завжди продуценти, створювачі органічної маси;
II трофічний рівень – рослиноїдні консументи (консументи I порядку);
Ш трофічний рівень – м'ясоїдні консументи, які живуть за рахунок рослиноїдних форм (консументи II порядку);
IV трофічний рівень – консументи, що живляться іншими м'ясоїдними (консументи Ш порядку).
Енергетичний баланс консументів: Р = П + Д + Н, де Р – раціон консумента, тобто кількість їжі, спожитої ним за певний період часу; П – продукція, тобто витрати на ріст; Д – витрати на дихання, тобто на підтримання обміну речовин за той же період, а Н – енергія незасвоєної їжі.
Основна частина спожитої з їжею енергії іде у тварин на підтримання їхньої життєдіяльності, і лише порівняно невелика – на побудову тіла, ріст і розмноження. З цього факту випливає дуже важливий висновок, а саме: більша частина енергії при переході з однієї ланки ланцюга живлення в іншу втрачається, тому що до наступного споживача може надійти лише та енергія, що міститься в масі з'їденого організму.
Отже, запас енергії, накопичений зеленими рослинами, в ланцюгах живлення швидко вичерпується. Тому ланцюги живлення не можуть включати в себе більше 4-5 ланок. Втрачена в ланцюгах живлення енергія може бути поповнена тільки надходженням нових її порцій. Ось чому в екосистемах не може бути кругообігу енергії, аналогічно до кругообігу речовин. Екосистема функціонує лише за рахунок спрямованого потоку енергії, постійного надходження її ззовні у вигляді сонячного випромінювання або готових запасів органічної речовини.
Трофічні ланцюги, які починаються з фотосинтезуючих організмів, називають ланцюгами виїдання (або пасовищними ланцюгами, або ланцюгами споживання), а ланцюги, що починаються з відмерлих решток рослин, трупів і екскрементів тварин – детритними ланцюгами розкладання. Таким чином, потік енергії, що входить в екосистему, розбивається далі мовби на два основних русла, надходячи до консументів через живі тканини рослин або через запаси мертвої органічної речовини, джерелом якої є фотосинтез.
Швидкість, з якою продуценти екосистеми фіксують сонячну енергію в хімічних зв'язках синтезованої органічної речовини, визначає продуктивність угруповань. Органічну масу, створювану рослинами за одиницю часу, називають первинною продукцією угруповання. Продукцію виражають кількісно сирою чи сухою масою рослин або в енергетичних одиницях – еквівалентному числі джоулів.
Валова первинна продукція – кількість речовини, створюваної рослинами за одиницю часу при даній швидкості фотосинтезу. Частина цієї продукції іде на підтримання життєдіяльності самих рослин (витрати на дихання). Частина створеної органічної маси, що лишилася, характеризує чисту первинну продукцію, яка являє собою величину приросту рослин. Чиста первинна продукція – це енергетичний резерв для консументів і редуцентів. Перетворюючись у ланцюгах живлення, вона йде на поповнення біомаси гетеротрофних організмів.
Приріст за одиницю часу маси консументів – це вторинна продукція угруповання. Вторинну продукцію обчислюють окремо для кожного трофічного рівня, оскільки приріст маси на кожному з них відбувається за рахунок енергії, яка надходить з попереднього.
Екосистеми дуже різноманітні щодо порівняння швидкості створення і витрачання як первинної продукції, так і чистої вторинної продукції на кожному трофічному рівні. Однак всім без винятку екосистемам властиві певні кількісні співвідношення первинної і вторинної продукцій, які дістали назву правила пірамід: на кожному попередньому трофічному рівні кількість біомаси, створюваної за одиницю часу, більша, ніж на наступному. Графічно це правило виражають у вигляді пірамід, які звужуються вгору і утворені поставленими один на одного прямокутниками однакової висоти, довжина яких аналогічна масштабам продукції на відповідних трофічних рівнях. Розрізняють 3 типи екологічних пірамід: біомас, чисел та продуктивності.


Пр – продуценти;
РК – рослиноїдні консументи;
МК – м'ясоїдні консументи;
ФП – фітопланктон;
ЗП – зоопланктон.
Піраміда продукції відображає також закони витрачення енергії у ланцюгах живлення. Тому, якщо піраміди чисел та біомас можуть мати різний (звичайний та перевернутий) вигляд в угрупованнях з певною трофічною структурою, то піраміда продукції має лише універсальний характер.
Екологічні фактори
Екологічні фактори (ЕФ) – всі складові (елементи) природного середовища, які впливають на існування й розвиток організмів і на які живі істоти реагують реакціями пристосування. Усю сукупність ЕФ поділяють на3 групи: абіотичні (неорганічні умови: хімічні й фізичні, такі як склад повітря, води, ґрунтів, температура, світло, вологість, радіація, тиск тощо); біотичні (форми взаємодії між організмами, хазяїн-паразит) та антропогенні (вплив людської діяльності).
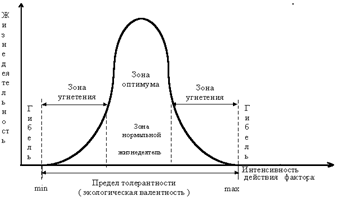
Лімітуючим фактором може виступати не лише недостатність, але і надлишок будь-якого ЕФ, наприклад, загибель врожаю від частих дощів, перенасичення ґрунтів добривами і т.д. (закон толерантності–Шелфорда).
 Широку екологічну валентність виду по відношенню до абіотичних факторів середовища позначають, додаючи до назви цього фактора приставку „еври”. Наприклад, тварини, які здатні переносити широкі коливання:
Широку екологічну валентність виду по відношенню до абіотичних факторів середовища позначають, додаючи до назви цього фактора приставку „еври”. Наприклад, тварини, які здатні переносити широкі коливання:
ü температури, називають еври т е р м н и м и;
ü солоності – еври г а л і н н и м и;
ü освітлення – еври ф о т н и м и;
ü будь-яких абіотичних факторів – еври б і о н т н и м и (таргани, колорадські жуки, вовки, миші, очерет, пирій).
Нездатність переносити значні коливання факторів або низька екологічна валентність позначається приставкою „стено”, наприклад тварини, які не здатні переносити широкі коливання:
ü температури, називають стено т е р м н и м и;
ü солоності – стено г а л і н н и м и;
ü освітлення – стено ф о т н и м и;
ü будь-яких абіотичних факторів – стено б і о н т н и м и (орхідеї, форель, далекосхідний рябчик, глибоководні риби).
Оскільки у природі всі фактори діють сумісно, то оптимальна зона та межі витривалості організму по відношенню до певного фактору середовища можуть зміщатись в залежності від того, в якому сполученні діють одночасно інші фактори (закономірність взаємодії та компенсації екологічних факторів). Однак взаємна компенсація має певні межі і повністю замінити один із факторів іншим неможливо. Отже, у сі умови середовища, необхідні для підтримки життєдіяльності, є рівнозначними для живого організму, будь-який фактор може обмежувати можливості існування організмів (закон рівнозначності усіх умов життя).
Екологічна ніша – це загальна сума всіх вимог організму до умов існування, включаючи простір, який він займає, функціональну роль в товаристві (наприклад, трофічний статус) та його толерантність відносно факторів середовища – температури, вологості, кислотності, складу ґрунту тощо.
Три критерії визначення екологічної ніші можна визначити як просторову, трофічну та багатовимірну.
Просторова ("адреса" проживання) – місцезнаходження організму у просторі.
Трофічна ніша ("професія" організму) – характеризує особливості живлення і, відповідно, роль організму у співтоваристві.
Багатовимірна ніша – це ділянка гіперпростору, вимірами якої є різні екологічні фактори.
Два види не можуть займати одну й ту ж саму екологічну нішу (принцип Гаузе).
біотичні екологічні фактори
| Тип взаємодії | Результати взаємодії для виду | Загальний характер взаємодії | |
| А | В | ||
| 1. Конкуренція | – | – | Взаємне пригнічення обох видів |
| 2. Хижацтво | + | – | Взаємодія сприятлива для популяції хижака А та несприятлива для популяції жертви В |
| 3. Паразитизм | + | – | Взаємодія сприятлива для популяції паразиту А та несприятлива для популяції хазяїна В |
| 4. Аменсалізм | – | Популяція В пригнічує популяцію А, але сама не отримує при цьому користі | |
| 5. Симбіоз (мутуалізм) | + | + | Взаємодія сприятлива для обох популяцій |
| 6. Коменсалізм | + | Популяція коменсала А отримує користь від об’єднання, а для популяції В це об’єднання байдуже | |
| 7. Нейтралізм | Жодна популяція не впливає на іншу |
Екологічна ніша як функціональне місце виду у системі життя не може також довго залишатися порожньою – про це говорить правило обов’язкового заповнення екологічних ніш: порожня екологічна ніша завжди природно заповнюється.
висновки:
1) відношення між живими організмами є одним з регуляторів чисельності та просторового розподілу організмів в природі;
2) негативна взаємодія між організмами проявляється лише на початкових стадіях розвитку співтовариства або при порушенні природних умов; у щойно сформованих або нових асоціаціях ймовірність виникнення сильних негативних відносин є більшою, ніж у старих асоціаціях;
3) в процесі еволюції та розвитку екосистем простежується тенденція до зменшення ролі негативних взаємовідносин більше, ніж у старих асоціаціях.
|
|
|
|
Дата добавления: 2014-01-11; Просмотров: 569; Нарушение авторских прав?; Мы поможем в написании вашей работы!