КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Цистеин
|
|
|
|
Пролин
Глицин
Серин
Аспарагин
Образование аспарагина из аспартата, катализируемое аспарагинсинтетазой, сходно с синтезом глутамина. Аспарагинсинтетаза млекопитающих в качестве источника азота использует не ион аммония, а глутамин и, следовательно, не «фиксирует» неорганического азота. Бактериальные же аспарагинсинтетазы используют ион аммония, следовательно, «фиксируют» неорганический азот. Как и в случае других реакций, сопровождающихся образованием РРi, последующий гидролиз РPi до Pi с участием неорганическойпирофосфатазы обеспечивает энергетически благоприятные условия для протекания реакции. Особенность: донором азота в данной реакции выступает α-аминогруппа глутамата (а не амидная группа), поэтому образуется амид α-кетоглутарата – глутарамат.
Важно! Выше рассмотренные реакции синтеза глутамата и аспартата носят обратимый характер (реакции дегидрогеназ и аминотрансфераз обратимы!), в то время как реакции синтеза глутамина и аспарагина (катализируются синтетазами) идут с высвобождением значительного количества энергии (распад ATP) и не обратимы.
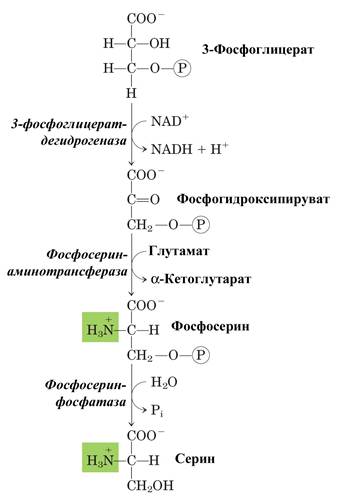
Серин образуется из промежуточного продукта гликолиза – 3-фосфоглицерата. В первой реакции α-гидроксильная группа при участии NAD+ окисляется в кетогруппу (фермент 3-фосфоглицератдегидрогеназа). Далее в результате переаминирования образуется фосфосерин (фосфосерин-аминотрансфераза), который затем дефосфорилируется, образуя серин (фосфатаза).
Также возможен вариант, когда сначала происходит дефосфорилирование 3-фосфоглицерара до глицерата (фосфатаза), а затем уже идут реакции окисления (глицератдегидрогеназа) и переаминирования (серин-аминотрансфераза).
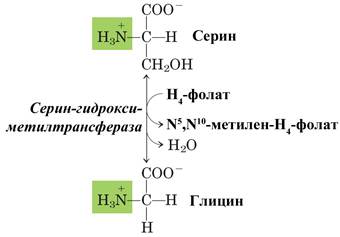
Другим важным путем образования серина является его обратимый синтез из глицина. Донором метильной группы является N5,N10-метилен-тетрагидрофолат, переходящий в тетрагидрофолат. Гидроксильная группа серина поступает из воды. Реакция катализируется серин-гидроксиметилтрансферазой.
А Б
Рис. А – синтез серина из 3-фосфоглицерата;
Б – реакция взаимопревращения серина и глицина.
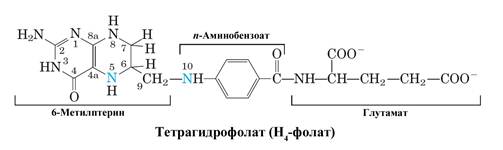
Рис. Структура тетрагидрофолата – одного из основных переносчиков одноуглеродных фрагментов в клетке (синим указаны атомы азота 5 и 10, к которым происходит присоединение одноуглеродных фрагментов – метильных, метиленовых, формильных и других групп).
Синтез глицина в тканях млекопитающих осуществляется несколькими путями.
В цитозоле печени содержится глицинтрансаминаза, катализирующая синтез глицина из глиоксилата, донором аминогруппы выступает глутамат (или аланин). В отличие от большинства реакций переаминирования равновесие этой реакций сильно смещено в направлении синтеза глицина.
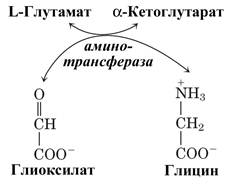 Рис. Синтез глицина из глиоксилата.
Рис. Синтез глицина из глиоксилата.
Два важных дополнительных пути, функционирующие у млекопитающих, используют для образования глицина серин (см. синтез серина) и холин.
|
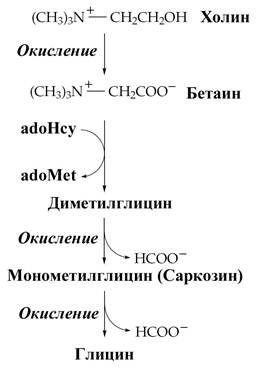
В клетках печени хордовых существует еще один путь синтеза глицина, катализируемый ферментом глицинсинтетазой. Глицин образуется из неорганических соединений: CO2 и NH4+. Донором дополнительного одноуглеродного фрагмента выступает N5,N10-метилен-тетрагидрофолат, восстановление происходит за счет энергии NADH.
У млекопитающих и некоторых других организмов пролин образуется из глутамата путем обращения реакций катаболизма пролина.
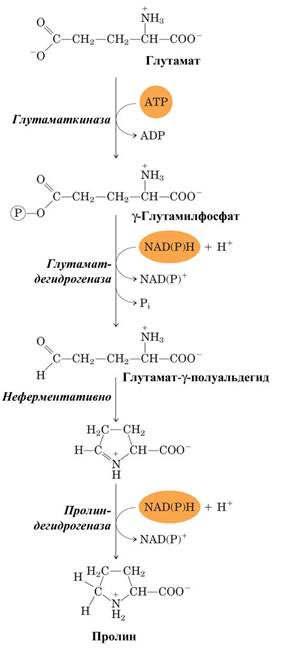
Рис. Синтез пролина.
Цистеин, относящийся к заменимым аминокислотам, образуется из незаменимого метионина и заменимого серина. Сначала происходит превращение активация метионина с образованием S-аденозилметионина (см. лекцию 3), затем метилтрансфераза переносит метильную группу с S-аденозилметионина на какой-либо акцептор (тетрагидрофолат, диметилглицин, фосфатидилэтаноламин) и образуется S-аденозилгомоцистеин. Данное соединение распадается на аденозин и гомоцистеин. Далее гомоцистеин взаимодействует с серином.
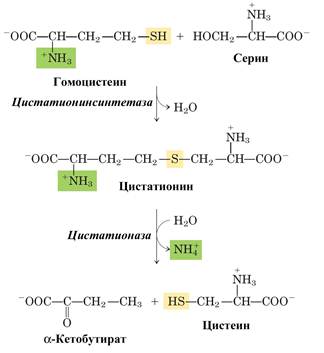
Получаемый в последней реакции α–кетобутират может подвергаться дальнейшему окислению с образованием пропионил-СоА.
|
|
|
|
Дата добавления: 2014-01-14; Просмотров: 965; Нарушение авторских прав?; Мы поможем в написании вашей работы!