КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Репарация двунитевых разрывов ДНК путем негомологического воссоединения концов (NHEJ)
|
|
|
|
Как мы уже отмечали, главной задачей систем репарации DSB является необходимость восстановления непрерывности поврежденной хромосомы, то есть сшивания именно тех двух концов ДНК, которые появились вследствие одного и того же DSB. Для этого их нужно быстро фиксировать в ядре и изолировать от возможного взаимодействия с концами, образованными другими DSBs, не мешая при этом правильному процессингу концов ДНК и их сшиванию лигазой. Важно понимать, что появление столь серьезного повреждения ДНК как образование DSB, часто сопровождается сопутствующими повреждениями азотистых оснований и сахарофосфатного каркаса ДНК. Такие повреждения должны быть удалены с концов ДНК с помощью нуклеаз до их сшивания ДНК-лигазами. Из этого следует, что при NHEJ в местах воссоединений почти всегда (кроме редких случаев образования «чистых» двойных разрывов, не требующих нуклеазной расчистки) образуются микроделеции, то есть такая репарация является по самой своей природе неточной. Но геномы высших эукариот в основном (более чем на 90 %) состоят из некодирующих последовательностей, и неточность NHEJ редко приводит к вредным мутациям, хотя и может вносить свой вклад в микроделеционный полиморфизм ДНК. Кроме того, нужно иметь в виду, что клетка способна эффективно репарировать лишь небольшое число DSB (несколько десятков), и количество микроделеций, возникающих при NHEJ, относительно невелико.
В принципе при воссоединении «неправильных» концов ДНК репарация DSB путем NHEJ может приводить и к большим делециям и транслокациям. Вероятнее всего, это происходит при большом количестве DSB, то есть у репарации путем негомологического воссоединения концов есть какая-то своя система отбора и сравнения тех концов, которые должны быть объединены для «правильного» воссоединения, но при избытке повреждений она с этой задачей не справляется.
На рис. 23 видно, что первым к месту разрыва подходит гетеродимер Ku, состоящий из прочно связанных друг с другом белков Ku70 (70кд) и Ku80(86кд). Он образует ассиметричное кольцо, которое надевается на свободный конец ДНК, как гайка на болт, цепляется за ДНК, и может продвигаться в глубину, позволяя связываться с длинным ДНК-концом другим белкам.
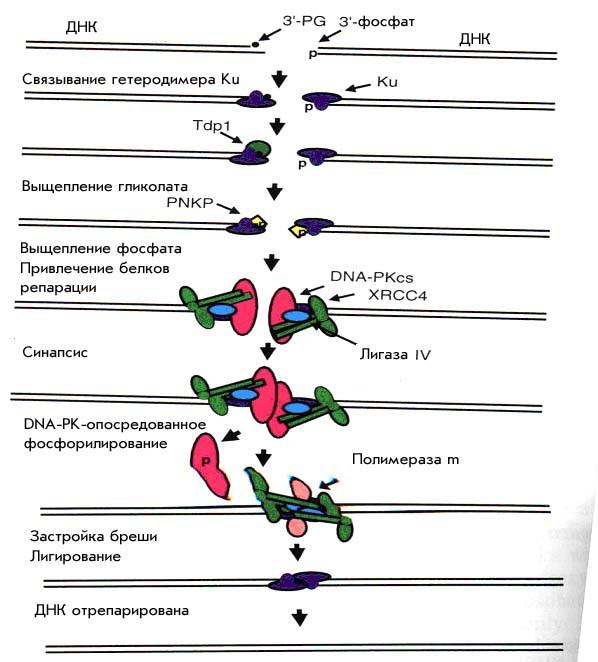
Рисунок 23. Репарация путем негомологического воссоединения концов (NHEJ)
Обычно гетеродимер Ku покрывает 16–18 н.п. Вероятно, именно два гетеролимера Ku, связавшиеся с разными концами одного и того же DSB, удерживают эти концы вместе на начальной стадии репаративной реакции и способствуют их «правильному» воссоединению.
Затем к комплексу присоединяется каталитическая субъединица протеинкиназы DNA-PKcs. Это белок размером 350Кд, обладающий протеинкиназной активностью, также имеющий открытый канал, в котором может быть связано 12 н.п. двойной спирали ДНК. По некоторым данным, она присоединяется к ДНК после Ku и сдвигает гетеродимер вглубь от конца молекулы ДНК. По другим данным, DNA-PKcs имеет четкую границу связывания с ДНК на уровне 30 н.п. от конца. Вероятнее всего, происходит и то и другое – этот белок «перекрывает» собой гетеродимер Ku. Образование комплекса Ku с DNA-PKcs необходимо для включения киназной активности DNA-PK. Собственно так и образуется особый гетеротример – ДНК-зависимая протеинкиназа. Критичным для ее активации является денатурация концов ДНК, с которыми связывается DNA-PKcs, и вовлечение нескольких нуклеотидов однонитевого фрагмента в канал ее голофермента. Активированная DNA-PK способна фосфорелировать большую группу белков, включая сам гетеродимер Ku и нуклеазу Artemis.
Активированная DNA-PKcs также способна автофосфорелироватся, что приводит к ее диссоциации от ДНК. У мышей мутации в гене, кодирующем DNA-PKcs, приводят к появлению животных с фенотипом SCID (several cobine immunodeficiency), иначе называемых «голыми» мышами.
Белок XRCC4 был выделен как фактор, ответственный за радиочувствительность и дефект V(D)J рекомбинации в клеточной линии китайского хомячка ХR-1. Этот белок имеет круглую «голову» и длинный (120Å) спиралевидный хвост. Обычно он образует гомотетрамер, в котором с каждого конца лежат по две «головы», соединенные четырьмя взаимодействующими хвостами. В репаративных реакциях XRCC4 всегда образует прочное соединение с лигазой IV, которая связывается с ним примерно в середине спирального хвоста. Эти белки формируют сложный комплекс, усиливающий ферментативную активность (например, лигирование) каждого из его участников. При этом кажется вероятным, что белок XRCC4 играет в этом комплексе структурирующую роль, так как одновременно он связывается и с белками KU и с DNA-PKcs.
Нуклеазами, участвующими в NHEJ, являются WRN (Werner syndrome nuclease), Artemis и MRE11 (meiotic recombination exonuclease, дрожжевой гомолог называется RAD58). MRE11 и WRN обладают 3’-5’ экзонуклеазной активностью и «предпочитают» протяженные 3’-концы, при этом гетеродимер Ku активирует активность WRN, а DNA-PK ее подавляет. В тоже время WRN обладает и ДНК-геликазной активностью, важной для процесса репарации с помощью гомологической рекомбинации (HR). Artemis обладает 5’-3’ экзонуклеазной активностью, но после фосфорелирования DNA-PK приобретает еще и эндонуклеазную, которая позволяет ему расплетать шпилечные петли, убирая выступающие 5’-концы и укорачивая 3’. Мутации, инактивирующие этот белок приводят у человека к крайне тяжелому наследственному синдрому радиочувствительности, сопряженной с иммунодефицитом (RS-SCID syndrome), подобному SCID у мышей.
Образование DSB после ионизирующего излучения почти всегда сопровождается фрагментацией ДНК при участии свободных радикалов, и позиционное расхождение между разрывами на отдельных нитях составляет не менее двух нуклеотидов. Это приводит к появлению выступающих частично комплементарных концов и однонуклеотидной бреши на каждой нити. Разрывы обычно имеют 5’-фосфатный (гидроксильный) конец, в то время как 3’-конец блокируется фосфатными или дезоксирибозными фрагментами и обычно представляет собой 3’-фосфогликолат (—PO4CH2COOH; PG). То есть для успешной репарации предварительно необходимо убрать блокирующую 3’-конец PG группу. Процесс NHEJ с этим успешно справляется, но именно по этой причине он обычно сопровождается микроделециями. И только тупые концы могут воссоединяться практически без вырезания концевых нуклеотидов.
Ферменты, способные убирать 3’-PGs идентифицированы. Это апуриновая/апиримидиновая эндонуклеаза APE1, а также белки TDP1 и PNKP, которая не только «приводит в порядок» 3’-конец, но в тоже время может фосфорелировать 5’-гидроксильные концы, причем это фосфорелирование одновременно проходит на обеих нитях ДНК. Данная реакция происходит в первые же минуты и не нуждается в полностью собранном репарационном комплексе. Более того, присоединение DNA-PKcs блокирует реакцию вымывания 3’-PGs, что заставляет нас предположить наличие фермента, сходного по действию с ранее описанными, но способного работать в контексте всего репарационного комплекса. После ограниченного расплетания двунитевой ДНК в месте разрыва и «зачистки» на образующихся концах происходит их сопоставление по участкам микрогомологии (1–3 нуклеотида).
Зашивание бреши ведет ДНК-полимераза μ (Polμ), одна из полимераз семейства Х, включающего ДНК-полимеразы μ, β и λ. Главное их различие между собой состоит в том, что полимераза μ образует прочный комплекс с XRCC4/LigIV, а полимераза β на это не способна. Polμ имеет домен, крайне сходный с таковым терминальной трансферазы, которая способна к добавлению нуклеотида к цепи при отсутствии матрицы в процессе V(D)J рекомбинации. То есть оба эти фермента вовлечены сходным образом в белковые комплексы, ведущие воссоединение концов, но в различных ситуациях. При этом клетки, дефектные по гену polμ не являются гамма-чувствительными, что говорит о том, что есть и другая полимераза, вероятнее всего – Polλ, способная полноценно замещать Polμ в реакции застройки брешей, так именно она (как и Polμ), а не репарационная полимераза β, имеет С-мотив, сходный с таковым белка BRCA1 (BRCT-motif). Этот мотив необходим для взаимодействия полимеразы с XRCC4 с образованием единого комплекса KU/Polμ/XRCC4/LigIV. Представлятся вероятным, что гетеродимер Ku остается связанным с ДНК вплоть до завершения процесса лигирования.
DNA-PKcs участвует в двух основных реакциях – во-первых, она способна удерживать рядом концы двунитевого разрыва, играет в этом удержании главную роль и делает это значительно лучше, чем гетеродимер Ku. Во-вторых, она способна к автофосфорелированию. Причем у нее есть по меньшей мере 5 сайтов автофосфорелирования, и это автофосфорелирование крайне функционально значимо. (Мутации хотя бы в одном из 5 известных сайтов приводят к повышению радиочувствительности). Вероятно, таким образом модулируется активность DNA-PK во время различных стадий репарационной реакции. К тому же DNA-PK фосфорелирует и другие белки репаративного комплекса. Складывается представление, что она связывается с концами ДНК и удерживает их до тех пор, пока не будет сформирован весь репаративный комплекс и не будут найдены «партнеры» по объединению. В тот момент, когда две молекулы DNA-PKcs от разных частей DSB вступают в контакт, они взаимно фосфорелируют друг друга и диссоциируют от репарационного комплекса, или, по крайней мере, отодвигаются от места реакции. В-третьих, DNA-PK может быть необходима для фосфорилирования нуклеазы Artemis, то есть для активации ее способности открывать шпильки во время V(D)J рекомбинации и вырезания выступающих концов во время репарационной реакции NHEJ.
На рис. 23 нужно особо отметить, что комплекс XRCC4/LigIV привлекается в репарационную реакцию еще до того, как Polμ проведет репаративный синтез. Это свидетельствует о необходимости структурного взаимодействия между Polμ и XRCC4 и о вовлечении комплекса XRCC4/LigIV в сам процесс заполнения бреши. Гетеродимер Ku выравнивает выступающие концы ДНК обеих молекул, а глобулярные «головы» белка XRCC4 связываются благодаря своему ДНК-связывающему мотиву несколько дистальнее Ku, и таким образом определяют положение лигазы IV, которая свяжется с центральной частью С-концевого хвоста XRCC4 для последующего лигирования концов. На каждой нити работает тример, состоящий из 2 молекул XRCC4 и одной LigIV. Причем этот тример образуется до того, как DNA-PKcs диссоциирует от репарационного комплекса. Сами DNA-PKcs также прочно связаны с С-хвостами XRCC4 на разных сторонах разрыва, и при их синапсисе, взаимном автофосфорилировании и диссоциации от ДНК четыре молекулы XRCC4 образуют тетрамер, сплетаясь хвостами. Последним на отрепарированной ДНК остаются два объединившихся (или расположенных прямо друг за другом) гетеродимера Ku.
Мы описали первым именно этот процесс репарации двунитевых разрывов потому, что, не нуждаясь в партнере для рекомбинации, он может протекать в любой фазе клеточного цикла, и к тому же используется клеткой в 1500 раз чаще, чем HRR.
|
|
|
|
|
Дата добавления: 2014-11-06; Просмотров: 938; Нарушение авторских прав?; Мы поможем в написании вашей работы!