КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Иллюстративті материал
|
|
|
|
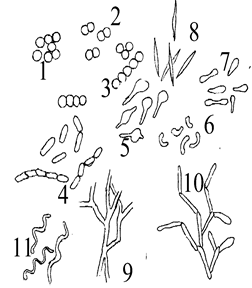
Сурет 1. Бактериялардың негізгі пішіндері:
|
Бактериялардың негізгі пішіндері: 1-3. Кокктар (стафилококктар, диплококктар, стрептококктар. 4. Таяқшалар (стрептобациллалар). 5. Клостридиялар. 6. Вибриондар. 7. Коринебактериялар. 8. Фузобактериялар. 9. Нокардиялар (шынайы тармақтану). 10. Актиномицеттер (жалған тармақтану). 11. Спирохеталар.
Сурет 2. Бактериялардың морфологиясы бойынша жіктелуі:
| стафилококктар |
 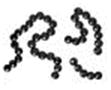
| стрептококктар |

| сарцина |

| диплококктар |
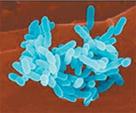 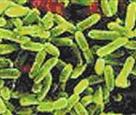 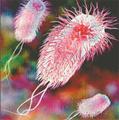
  
| бактериялар (спора түзбейтін таяқшалар) |

| Клостридиялар (анаэробты спора түзетін микробтар) |

| Спириллалар |
Сурет 3. Имектелген пішінді бактерияла р
|
| Спирохеталар |
Бақылау сұрақтары (кері байланыс)
1. Өмірдің негізгі пайда болу кезеңдері.
2. Микроорганизмдердің даму және пайда болу эволюциясы.
3. Микроорганизмдер қайда қолданады?
4. Өмірдің негізгі пайда болу кезеңдері.
5. Бактерияның L-трансформациясы дегеніміз не?
Тақырыбы: «Бактериялардың физиологиясы және биохимиясы»
. Микроорганизмдерді дақылдандыру үшін арнайы анаэробты жағдайды жасау әдістері.
1. Физикалық – арнайы оттегісіз қоспа қосылған ауаны ығыстыру (көбіне- N,- 85%, СО2
10%, Н2 - 5%).
2. Химиялық - оттегіні химиялық жұтушылар қолданады.
3.Биологиялық - қатал аэробтар мен анаэробтарды бірге дақылдандыру (аэробтар оттегіні жұтып, анаэробтардың өсуіне қолайлы жағдай жасайды).
4.Аралас – бірнеше түрлі әдістер қолданады.
Айта кететін жәйт, қатал анаэробтарға оптимальді жағдай жасау өте қиын іс.Сол сияқты дақылдандыруда оттегісіз жағдайды тұрақты қамтамасыз ету мүмкін емес, сол себепті ерітілген оттегі құрамынсыз арнайы орталар, оларды алу, жеткізу, анаэробтық жағдайда материалды егу аса қажет.
Оның бірнеше әдісі бар, яғни анаэробтардың өсуіне келетін жағдаймен қамтамасыз ету үшін – қоректік орталарды алдын-ала қайнатып алу, терең агар бағанасына егу, оттегінің түсуін азайтатын орталарды вазелин майымен көмкеру, герметикалық жабылатын флакондар мен пробиркаларды, шприцтерді және инертті газы бар зертханалық ыдыстарды, жанып тұрған май шамы бар тығыз жабылатын эксикатор қолдану. Анаэробты жағдай туғызу үшін арнайы приборлар анаэростаттар қолданады. Бірақ қазіргі уақытта анаэробты және микроаэрофильді жағдай жасау үшін қарапайым және өте тиімді жүйе «Таз-пак»'қолданады, оның арнайы герметикалық жабық ыдыстардағы атмосфералық ауаны, газ қоспаларымен ығыстыру принципі бойынша жұмыс жасайтын арнайы газорегенерирлеуші пакеттері бар.
4. Қоректік орталарда микроорганизмдерді дақылдандыру принциптері
1. Микробтарға сәйкес келетін қоректік компоненттерінің барлығын барынша қолдану.
2. Оптимальді температура, рН, гН2, иондардың концентрациясы,оттегімен қанығу деңгейі, газдық құрамы және қысымы.
Микроорганизмдерді оптимальді температурада термостатта қоректік орталарға дақылдандырады, себебі бұл жағдай инкубациялауды қамтамасыз етеді.
Температуралық оптимум бойынша микроорганизмдерді басты үш топқа бөледі.
1.Психрофилдер - +20 градус Цельсиядан төмен температурада өседі.
2. Мезофиллдер - 20-дан 45 градус температуралық диапозонда өседі (көбіне оптимум- 37 градус С).
3.Термофилдер - 45 градустан жоғары температурада өседі.
5. Қоректік ортаға қысқаша сипаттама
Тығыздығы бойынша сұйық,тығыз (1,5-3% агар) және жартылай сұйық (0,3-0,7 % агар) орталар деп жіктейді.
Агар – теңіз өсінділерінен алынған күрделі құрамды полисахарид,тығыз (қатты) орталарға арналған. Көміртегі және азоттың универсальді көзі ретінде пептондар-азықтарды пепсинмен белоктық ферментациялау қолданады, түрлі гидролизаттар- етті, балықты, казеинді, ашытқылы және т.б..
Тағайындалуы бойынша орталарды келесі топтарға жіктейді
-универсальді (қарапайым), пригодные для различных нетребовательныхмикроорганизмов (етті- пептонды сорпа- ЕПС. етті- пептонды агар- ЕПА);
- арнайы- универсальді орталарда өспейтін микроорганизмдер үшін (Мак-Коян ортасы туляремия қоздырғышы үшін, Левенштейн- Иенсен ортасы туберкулез қоздырғышы үшін);
- дифференциальді-диагностикалық орта- микроорганизмдерді ферментативті белсенділігі және дақылдық қасиеттері бойынша дифференциациялау үшін (Эндо, Плоскирев, Левин, Гисса орталары);
- селективті (элективті)- микроорганизмдердің нақты бір түрлерін бөліп алу және өсуін тежеу үшін – пептонды су,селенитті орта, Мюллер ортасы. үшіндля выделения определенных видов микроорганизмов и подавления роста сопутствующих- пептонная вода, селенитовая среда, среда Мюллера.
Шығу тегі бойынша табиғи,жартылай синтетикалық және синтетикалық деп бөледі.
Бактериялардың тығыз және сұйық қоректік ортада өсу сипаты: толығымен өсуі,колония,тұнба, пленка түзу, лайлануы.
Таза дақыл - микроорганизмдердің бір түрінің популяциясы.
Бактерия ферменттері экзо- және эндоферменттерге бөлінеді.
Экзоферменттер сыртқы ортаға бөлінеді, жоғарымолекулярлы органикалық байланыс үрдісін ыдыратуды жүзеге асырады. Экзоферменттердің түзілу қабілеті көбіне бактерия инвазивтілігін анықтайды, яғни кілегей қабаты, дәнекер тіні және басқа тіндік барьерлерден өту қабілеттілігі. Мысалы: гиалуронидаза гиадурон қышқылын ыдыратады, ол клетка аралық зат құрамына кіреді, тіндер өткізгіштігін жоғарлатады (клостридиялар, стрептококктар, стафилококктар және көптеген микроорганизмдер); нейраминидаза кілегей қабатынан өтіп кетуді жеңілдетеді де, клетка ішіне еніп, клеткааралық кеңістікке таралады (тырысқақ вибрионы, күл таяқшасы, тұмау вирусы және басқалары). Осы топқа антибиотиктерді ыдырататын энзимдер жатады.
Бактериологияда микроорганизмдерді биохимиясы бойынша танып-білу үшін ферменттер әсерінің нәтижесі мен соңғы өнімдердің алатын орны аса зор. Осыған сәйкес ферменттердің микробиологиялық (жұмысшы) жіктелуі.
1.Сахаролитикалық.
2. Протеолитикалық.
3. Аутолитикалық.
4. Тотығу-тотықсыздану.
5. Патогенділік ферменттері (вируленттілігі).
Клетканың ферменттік құрамы геноммен анықталады және тұрақты белгісі болады. Микроорганизмдердің биохимиялық қасиетін білу олардың ферменттік жиынтығын танып-білуге септігін тигізеді. Көміртегі мен белоктардың басты ферменттейтін өнімдері- қышқыл, газ, индол, күкіртсутегі, дегенмен микроорганизмдердің түрлі спектрлері одан да көлемдірек. Вируленттіліктің басты ферменттері - гиалуронидаза, плазмокоагулаза, лецитиназа. нейраминидаза, ДНК-аза. Патогенділік ферменттерін анықтау көптеген микроорганизмдерді бөліп алу мен танып-білуінің патологияда маңызы зор.
Микроорганизмдердің кейбір ферменттері медицина мен биологияда әр-түрлі заттар алу үшін (аутолитикалық, протеолитикалық), гендік инженерияда (рестриктазалар, лигазалар) кеңінен қолдану.
Бақылау сұрақтары (кері байланыс)
1.Микроорганизмдердің ішкі ортамен өзара байланысы қалай іске асады?
2.Тірі ағзалардың тігінен және көлденең эволюциялары қалай жүреді?
3.Қандай бактериальді тасымалдау жүйесін білесіз?
4.Микроорганизмдерді ішкі ортада не синтездеп, секреттейді?
Тақырыбы: Жалпы вирусология. Вирустарды жүйелеу негіздері. Вириондардың құрылымы және химиялық құрамы. Бактериофагтар.
Дәрісті оқу мақсаты: студенттерді вирустар мен бактериофагтың құрылысындағы ерекшеліктермен таныстыру, вирустың торша-иесімен әрекеттесу кезеңдері және осының нәтижесі, вирустарды дақылдандыру әдістері, бактериофагты тәжірибеде қолдану, сонымен қатар хромосомадан тыс генетикалық ақпаратты тасымалдаушылар, медициналық биотехнология.
ВИРУСТАР ЖӘНЕ ОЛАРДЫҢ ЖІКТЕЛУІ, ҚҰРЫЛЫСЫ,
РЕПРОДУКЦИЯСЫ, ӨСІРУ ӘДІСТЕРІ. ВИРУСҚА ҚАРСЫ ИММУНИТЕТ.
Вирустар – тұқым қуалаушылық қасиеті бар, өзгеруге, көбеюге бейім өте ұсақ, тірі микроорганизмдер. Вирустардың бактериялардан айырмашылығы – клеткалық құрылысы болмайды, тек қана бір нуклеин қышқылы – ДНҚ немесе РНҚ болады. Клетка ішінде генетикалық дәрежедегі паразиттер болып саналады. Белок синтездейтін рибосомалары болмайды, сондықтанда оларда зат алмасу жүрмейді.
Вирустардың пайда болуы жөніндегі әртүрлі гипотезалардың арасында көп таралған пікір мынау: клеткалық ДНҚ-сынан ДНҚ вирустар, клеткалық РНҚ-дан РНҚ-вирустар пайда болады. Бұл гипотезаның негізгі дәлелі – клеткалық және вирустық нуклеин қышқылдарының ұқсастығы болып табылады.
ЖІКТЕЛУІ
Vira дүниесі екі үлкен топқа бөлінеді – ДНҚ және РНҚ – құратын топтар. Топтар тұқымдастықтың тармағынан, туыстардан және түрлерден тұрады. Жіктелу принципі мына белгілерге негізделген:
А. Вирустардың қасиеттері – нуклеин қышқылдары, капсид симметриясының типі, суперкапсидтің болуы және болмауы, вирионның бетіндегі рецепторлық құрылыстардың өзгешелігі, вирионның формасы және өлшемі, оның антигендік сипаттамасы:
Б. Сезімтал өзгерулердің тобы, әртүрлі органдар мен тіндерде тропизмі болады.
В. Инфекцияның әсер ету жолы және географиялық таралуы.
ҚҰРЫЛЫСЫ.
Вирус құрылысының компоненттері: капсомер-белокты бөлек бірлігі, капсид-капсомерлерден құралған, нуклеокапсид – нуклеин қышқылы мен капсид белогының комплексі, вирион – вирустың бүтін бөлшегі.
Вирустардың құрамы өте қарапайым. Олар нуклеин қышқылынан және оларды қоршап жататын – капсид және суперкапсид деп аталатын қабаттардан, сондай-ақ вирус қабығының сыртындағы рецепторлардан (синонимі – бекітуші белоктар, тікенектер) құралады. Аздаған ферменттер болады, олардың біреулері вирус нуклеин қышқылдарының репликациясына және вирус белоктарының транскрипциясына қажет, албасқалары – клетканың ішіндегі метоболиттік процесстерді өзгертіп немесе тежей отырып, зат алмасу процесстерінің вирустардың бөлшектерін құрастыру үшін жұмыс істетеді.
Морфологиялық құрылысы бойынша вирустар 2 топқа бөлінеді:
- жай, қарапайым құрылысы – нуклеин қышқылы және оны қоршаған капсидтен тұрады (полимиелит вирусы, аденовирустар, құтыру вирусы)
- күрделі құрылысты – нуклеин қышқылынан, капсид және суперкапсидтен тұрады.
ДНҚ немесе РНҚ вирионның ортасында орналасады. Вирус геномы 3-4-тен (парвовирустар) 150-ге дейін (шешек вирусы) геннен құралады. Әртүрлі вирустардың геномындағы нуклеин қышқылы бір немесе екі жіпшелі, үздіксіз немесе үзікті, түзу сызықты немесе сақиналы болуы мүмкін. Мысалы: ЖИТС вирусында екі үзіксіз түзу сызықты РНҚ жіптері, әрі 1/3 қысқартылған ішкі жіп; тұмау вирусында бір үзікті РНҚ жібі бар. Нуклеин қышқылы геномды белоктармен байланыстырады.
Капсид – жеке-жеке белоктық суббірліктен құралған – капсомерлерден тұрады. Капсомерлердің кеңістіктегі орнына сәйкес капсид симметриясының екі түрі болады – спиральдық және кубтық. Спиральды типінде капсид нуклеин қышқылын орап жатады, кубты типінде – капсид ортасында нуклеин қышқылы орналасқан қуыс дене тәрізді болады. Бактерия вирустарында – бактериофагтарда симметрияның кубтық болса, ал өсіндісінде – спиральдық болып келеді.
Құрылысы күрделі вирустардың суперкапсиді липопротеидтық комплекстен тұрады. Вирионның құрамындағы липидтер мен көмірсутектердің шығу тегі клеткалық екендігі осы күні мойындалып отыр.
Вирионның бетіндегі рецепторлардың формасы әртүрлі: таяқшалы, саңырауқұлақты, шоқпарлы т.б. Химиялық құрамы бойынша бұлар гликопротеидтердің қатарына жатады. Вирустар рецепторлар арқылы сезімтал клетканың бетіне жабысады (адсорбцияланады), ал сондай-ақ рецепторлардың белгілі физика-химиялық қасиеттері бар. Мысалы, тұмау вирусының 2 түрлі рецепторы бар – таяқшалық – гемагглютининдер және шоқпарлық – нейраминидазалар.
Гемагглютининдердің рецепторлық қызметінен басқа эритроциттерді гемолиздеу қабілеті бар, сонымен қатар ауру адамның организмінде талғамды антиденелер түзіліп, жоғары иммуногендік қасиет көрсетеді, талғамды антиденелер әсіресе рецепторлық құрылымдарға қарсы пайда болатындығына назар аудара кету керек. Вирустық талғамдылығы бар белоктар құрылысты және құрылыссыз түрге бөлінеді. Құрылысты белоктарға капсидтің, суперкапсидтің, рецепторлық құрылымдардың белоктары, сонымен қатар нуклеин қышқылымен байланысқан геномдық белоктар жатады.
Құрылыссыз белоктар вирионды репродукцияға дайындайды және төменде көрсетілген ферменттерден тұрады: нуклеин қышқылының репликациясына және транскрипциясына қажетті полимеразалар, вирустық талғамдылығы бар белоктарды бөлуші – протеазалар және реттеуші белоктар – ферменттер.
Вирустар мөлшеріне байланысты төмендегідей топтарға бөлінеді:
1. ұсақ – 50 нм кем (полимиелит вирусы, А және В гепатиттервирустары)
2. орташа – 150 нм кем (құтыру вирусы, тұмау вирусы)
3. ірі – 400нм дейін (шешек вирусы)
Вирус бөлшектерінің өлшемін анықтау әдістері:
1. диаметрі белгілі микропорлары бар мембраналық фильтр арқылы сүзу:
2. ультрацентрифгалау кезінде вирус бөлшектерінің тұнбаға түсу жылдамдығымен;
3. электрондық микроскоп көмегімен;
4. вирус бөлшектері суспензиясынан өткен иондаушы радиация ағынын өлшеу арқылы анықталады.
РЕПРОДУКЦИЯ.
Вирустардың жануарлардан, өсімдіктерден және микробтардан айырмашылығы олар тек қана тірі сезімтал клетканың ішінде дисьюниктивті жолымен көбейеді. Вирустар өздерінің белоктерын және нуклеин қышқылын синтездеу үшін, клетканың құрылысын және зат алмасу процесін пайдалынылады. Вирус жұққан клетка өзінің гентикалық және рибосомдық аппаратымен вирусспецификалық нуклеин қышқылдарының және белоктарының түзілуіне мүмкіндік туғызады. Осы нуклеин қышқылдарынан, белоктардан капсидтер, суперкапсидтер және рецепторлық құрылымдар түзіледі. Сондықтан, вирустар генетикалық паразиттер болып саналады. Және де, вирус компоненттерінің синтезі бөлек-бөлек жүргізіледі. Нуклеин қышқылдары цитоплазмада немесе ядрода, ал вирус белоктары клеткалық рибосомаларда жиналады және содан кейін барып олар қосылып бүтін вирион құрылу процесі басталады. Осындай көбею немесе репродукция әдісі, дисьюниктивті немесе бөлектенген әдіс деп саналады.
Репродукцияның негізгі кезеңдері: сезімтал клетканың бетіне вирус бөлшектерінің адсорбцилануы, вирус клетканың ішіне енуі, оның шешінуі; транскрипция, трансляция және репликация; вирус бөлшектерінің жиналуы және клеткадан шығуы.
1. Сезімтал клетка – нысана (мишень) бетіндегі рецепторларға вирустың адсорбциясы. Макроорганизмнің әртүрлі тіндер вирустық инфекцияның тропизмімен анықталады. Мысалы: тұмау вирусы – тек қана тыныс алу жолының шырышты қабығының клеткаларына бекінеді, құтыру вирусы – нерв клеткаларына, ЖИТС вирусы – лимфоциттерге бекінеді. Тұмау вирусы гемагглютинин рецепторлары арқылы тыныс алу жолдарының юоғарға бөліміндегі эпителиальді клеткалардың бетіндегі комплементарлы рецепторлар арқылы байланысады. Егер де вирус клетка ішіне өте алмаса, нейраминидаза – вирус бетіндегі бекітуші белок – вирусты бұл клеткадан бөліп алып басқа клеткаға өтуіне мүмкіндік жасайды. Бір клетканың бетінде спецификалық рецепторларлың саны 104-105 болады.
2. Вирустық бөлшектердің клеткаға енуі энергияға тәуелді процесс және екі әдіспен іске асырылады:
А) пиноцитоз арқылы клетка қабығынан өтуі. Цитоплазмалық мембрана вирус бөлшегін қамтып алып клетканың ішіне өтеді, вирус содан кейін мембранадан ажырайды (осындай әдіспен құтыру, полимиелит вирустары енеді).
Ә) вирус қабығымен клеткалық мембрананың бірімен-бірі қосылу әдісімен енуі (құрылысы күрделі вирустарға тән – тұмау, ұшық вирустары). Әдетте, клетканың ішіне репродукция процесіне қажетті вирустық нуклеин қышқылы және вирустық фермент – белктары енеді.
Б) Клетканың мембранасына сәйкес, вирустардың шешінуі әртүрлі болуы мүмкін. Мысалі: бактериофагтар өз қабығынан микробтық клетканың бетіне босатыады, шешек вирусы суперкапсидті клетка мембранасында қалдырып, капсидпен клетканың ішіне босатылады, полимиелит вирусы клеткаға еніп ішінде өзінің жалғыз қабығы – капсидтен босатылады. Шешінудің мағынасы мынау: тек қана қабықтан босатылған вирус көбейе алады, инфекция процесі сонда ғана дамиды.
3. а) транскрипция – бұл гентикалық коді заңы бойынша вирустық РНҚ немесе ДНҚ көшіріп жазылуымен байланысты иРНҚ-ның пайда болуы.
б) трансляция – бұл иРНҚ-дағы гентикалық ақпаратты аминқышқылдарының спецификалық жүйелілігі түріне көшіру процесі және вирусспецификалық белоктардың синтезінің басталуы. Қазіргі кезде вирустық иРНҚ клетканың рибосомальді аппаратымен қалай әрекеттеседі және рибосомаларда вирусспецификалық белоктар қалай жұмыс істейтіндігі әлі белгісіз. Синтезделген вирустық белоктар өзінің қызметін, олардан протеаза ферменттері арқылы кейбір ерекше жерлерде пептидтер кесіліп тастағаннан кейін және глюкозилденген, ацетилденген,фосфорилденгеннен кейін ғана істей алады;
в) репликация – бұл вирус нуклеин қышқылы молекуласының (ДНҚ немесе РНҚ) синтезі. Вирусспецификалық белоктардың синтезі тек қана клетка рибосомаларда жүрсе, ал вирустың нуклеин қышқылының репликациясы ядрода (ұшық вирусы, аденовирустар) немесецитоплазмада (шешек, құтыру, полимиелит вирустары) жүреді.
4. Вирус бөлшектерінің жинақталуының екі түрі болады;
а) вириондардың жинақталуы (толық жетілуі) клетканың ішінде өтеді (аденовирустар, шешек, полимиелит вирустары);
б) вирионның жинақталуы клеткадан шығу мезгілінде аяқталады (тұмау вирусы, ретровирустар).
Вирустар бөлігінің жинақталу процесі клетка мембраналарында жүреді: ядро, цитоплазмалық, эндоплазмалық мембраналарында жүреді. Клетка ішінде вирустық бөлшектердің жинақталу орны – фабрикалар (вируспен зақымдалған клеткаларды зерттегенде анықталады және қосындылар деп аталады). Олар цитоплазмада (құтыру инфекциясы кезіндегі Бабеш-Негри денешіктері) немесе ядрода (ұшық инфекцияда) болуы мүмкін.
Вирус бөлшектерінің клеткадан шығу әдістері:
а) «жарылу» жолымен, кейде жиналған вирус бөлшектері бір мезгілде клеткадан шыққанда оның бұзылып жойылуына себеп болады. Бұл әдіс құрылыстары жай вирустарға тән (полимиелит вирусы, аденовирустар);
б) «бүршіктену» жолымен, кейде вирус бөлшектері аздаған мөлшерде клеткадан шығады. Бұл әдіс құрылысы күрделі вирустарға тән (ұшық, тұмау вирустары). Клетка ұзақ уақыт тіршілігін сақтайды.
Жоғарыда қарастырылған вирус репродукциясы негізінде жедел және созылмалы инфекциялар тудыратын вирустың клеткамен қарым-қатынасын көрсетеді. Бұл қарым-қатынас продуктивті тип болып саналады. Вирустың клеткамен қарым-қатынасының екіншісі интегративтік тип болады. Бұл типте репродукция жоқ, ал вирус гендері клетка ДНҚ құрылысына енеді. Бірақ клетканың вируспен қарым-қатынасы тіндік клетканың атиптік түріне айналу қаупін туғызуы мүмкін.
ВИРУСТЫ ӨСІРУ ТӘСІЛІ
1. Вирустардың өсуі мен бөлінуінің бірден-бір тәсілі лабораториялық жануарларды зақымдау болып саналады. Жаңа туған жануарлар жиі пайдаланылады, себебі олар сезімтал келеді. Тәжірибелік жануарларды сұрыптау кезінде вирустарға олардың сезімталдығы мен қабылдағыштығына көңіл аудару керек. Құтыру вирусымен тышқандар, үй қояндарызақымданады, ал, тұмау вирусымен тышқандар және күзендер зақымдалынады. Вирусы бар материалды жануарларға егу (тері асты, бұлшық етке, венаға, миға, мұрын қуысы арқылы) жолы жануарлардыңтүрі мен вирустардың тропизміне байланысты.
2. Сыртқы ортаның обьектілерінен және аурулардан алынған вирустарды өсіру үшін тауық эмбрионы кеңінен қолданылады және осы жолмен өсірілген вирустардан вакциналарды дайындау, диагноз қоюға диагностикумдарды дайындау, вирустардың түрін анықтау үшін қажет.
Адам және жануарларда зақым келтіретін, көптеген вирустар көп немесе аз мөлшерде тауық эмбрионында көбейе алатындығы анықталған. Эмбрионның тығыз сыртқы қабаты сыртқы ортадан түсетін микроорганизмдерден қорғайды, сондықтан вирустар стерильді жағдайда көбейеді. Вирустарды өсіру үшін 4-12 тәуліктік эмбриондар пайдаланылады. Мысалы: тұмау вирусын бөліп алу үшін 11 тәуліктік эмбрионды қолданылады.
Өмір сүру қабілеті бар эмбриондарды жұмысқа іріктеп алу үшін оларды овоскоппен зерттейді. Жарық өткен қабықтың ішінде эмбрион, хорионалантоистық қабықтың ауа қуысының шекарасы көрінеді. Өмір сүргіштік қабілеті бар эмбриондар жылжымалы, қабыршықтарының тамырлары қанмен толтырылған.
Вирусы бар материалды алантоистық қуысқа (тұмау, ұшық, шешек вирустары), амнион қуысына (тұмау вирусы), сары қапшыққа (құтыру, ұшық вирустары), хорион-алантоистық қабыршықтарға (тұмау, шешек вирустары) енгізіледі. Хорион-алантоистық қабықты вирустарды көбейту үшін қолдану кең тараған әдістердің бірі болып табылады. Оның негізгі себебі, көптеген вирустар осы қабықта көбеюі белгілі бір морфологиялық өзгерістерге түседі. Көбінесе төмпешік (бляшка) тәрізді ақшыл дақтар пайда болады.
3. Вирустарды торша өсіндісінде өсіру.
Вирустарды өсіру үшін клетка культурасы пайдаланылады. Көптеген вирустар өздері көбею үшін сезімталдығы жоғары кульдураларды таңдап алатын болғандықтан, клетка культурасы универсальды культура болып табылады. Адамның, тауықтың, қойдың, сиырдың теңіз шошқасының, ақ тышқандардың трипсинденген эмбриональдық клеткалары және ересек маймылдардың бүйрек клеткасы жиі қолданылады. Сондай-ақ қатерлі ісік клеткаларының культурасы пайдаланылады. Трипсиндеудің негізі, протеолитикалық ферменттердің (трипсин, хемотрипсин т.б.) әлсіз ерітінділері арқылы клетка аралық байланысты үзе отырып, клеткалардың өмір сүру қабілетін сақтап қалуы болады.
Клеткадағы вирустардың цитопатикалық әсерінің байқалуы зақымдалған клеткадағы вирустардың бар екендігін анықтау үшін өте қолайлы.
Клетка культурасы басқа тәсілдерге қарағанда, вирустардың өсуіне біркелкі жағдай туғызады, өйткені клеткалар бір тканьге жатады. Қасиеттері ұқсас және арасында антиденелер, ерекшелігі жоқ тежеушілер болмайды.
Клетка жүзгінің in viro тіршілігін сақтау үшін құрамы күрделі қоректік орта (Игла, 199 ортасы) және тұзды ерітінділер (Хенкса, Эрла ерітіндісі, Тироде сұйығы) қолданылады. Қоректік орта клетка культурасының өсуін және көбеюін қамтамассыз етеді, ал тұзды ерітінді клетканың тек қана тіршілігін сақтауға қажет. Бактериялармен ластанудан сақтау үшін (әсіресе микоплазмадан) өсу ортасына антибиотиктер қосылады.
Клеткалар культурасының төмендегідей жіктелуі қолданылады:
А) тіршілігін сақтайтын тіндердің культурасы
Б) өсуші тіндердің культурасы, олардың өзі:
1. айқындалған тіндер бөлігінің культурасы – тіннің ұсақталған бөлімі тауық плазмасымен араластырылады, пайда болған қою бөліктерді бекітеді.
2. суспензияланған клетка культурасы – клетка суспензиясы үнемі магниттелген араластырғышпен немесе барабанмен, болмаса арнайы реактормен араластырып тұрады.
3. Бір қабатты клетка культурасы. Клетка әсері алдында шынының бетіне бекітіледі (пробирканың, матрацтың қабырғаларына) содан кейін қалыңдығы бір клеткадай қабат құрал өседі, соның арқасында болып жатқан өзгерістерді байқауға болады. Бірыңғай қабат культурадан біркелкі өмір сүргіштігі жақсы көп дәрежелі клеткалар алуға болады.
Бір қабатты клетка культурасы төмендегідей топтарға бөлінеді:
А) дифференцияланған клеткалар, олар 5-10 рет егуге бейім – бірінші реттік клетка культурасы:
Б) бөліне беретін клетка культурасы, атиптік клеткалардан алынады. Бұл клеткалар ағзадан тыс көбейеді HeLa, Hep-2, KB. Бөлінгіш атиптік клеткалар культурасы вакцин дайындау үшін вирустар жиынтығына қолданылмайды.
В) жартылай бөлінгіш немесе диплоидты клетка культурасы 50 пассажға дейін ұстай алады. Адам эмбрионының әртүрлі тіндерінен алынады. Диплоидты клеткаларды вирустардың өсуі үшін және вакциналарды дайындау үшін жиі қолданылады.
ВИРУСТАРҒА ҚАРСЫ ИММУНИТЕТ
Макроорганизмдегі қорғаныштық иммундық жауап вирусспецификалық антигендерге дамиды. Антигендерге структуралы және структуралы емес вирус белоктары және олардың нуклеин қышқылдарымен көмірсутегімен және липидтер комплекстері жатады. Вирусты антигендер беткейлік немесе түрлі спецификалық және тереңдегі немесе топты спецификалық болып бөлінеді. Мысалы: тұмау вирусының гемагглютининдері мен нейраминидазалары түрлі спецификалық антигендерге жатады, ал нуклеокапсидтер – топты спецификалыққа, ЖИТС вирусының беткейлік антигендері др 120 және др 41 белоктарымен, сондай-ақ Т4 рецептордың структуралық компоненттерімен көрсетілген. Ал, тереңдік антигендер ішкі белоктық қабатындағы р12 және р24 белоктарымен көрсетілген.
Иммундық жауаптың күші мен ерекшелігі көбінесе түрлі-спецификалық антигендердің қасиетімен байланысты. Мысалы: тұмау вирусының гемагглютининдері бір жағынан нейтралдайтын антиденелердің синтезіне себеп болады, екінші жағынан олар цитотоксикалық Т-лимфоцит үшін зақымдалған клетканың бетіндегі нысана ретінде саналады.
Вирустарға қарсы иммунитет гуморальды және клеткалы иммундық жауаптан, сондай-ақ спецификалық емес қорғаныш факторлардан құралады. Вирусты инфекцияларда гуморальдық иммунитет вирус нейтралдаушы антиденелер пайда болуымен байқалады. Қандағы антиденелердің мөлшері көптеген вирустық инфекцияның резистенттілігінің көрсеткіші немесе иммундық қорғаныштық қаншалықты жоғары екендігінің көрсеткіші болып бағаланады (қызылша, полимиелит, эпидемиялық паротит инфекциялары). Ұтымдылығы төмендегі көрсетілген себептерге байланысты:
А) антиденелер вирустардың рецепторларымен байланысады және осы байланыс арқылы сезімтал клетка рецепторларына вирустардың адсорбциялануын тоқтатады:
Б) антиденелердің липопротеидті қабаты бар вирус бөлшектерімен комплементтердің қатысуына әрекеттесуі және вирустардың ыдырауына әкеп соқтырады:
В) вирустық бөлшектердің Ig C қатарына жататын антиденелермен қосылуы олардың мононуклеарлы клеткаларымен және полиморфты лейкоциттермен фагоцитозына әкеп соғады. Ig C, М кластарына жататын вируснейтралдаушы антиденелер қанның сарысуында ұтымды, ал Ig А класына жататын антиденелер – шырышты қабатта тиімді.
Г) антиденелер вирустық бөлшектердің нейтралдауынан басқа жойылуымен аггрегациясына мүмкіндік береді.
Иммундық қорғаныштың клеткалық формасы вирусты инфекцияларда басты орын алады. Зақымданған клетканың қабығына вирустар өздерінің белоктарын орналастырады, сонан соң зақымданған клеткалар иммундық қорғаныштық клеткалық механизмі туғызатын ыдырау процесіне сезімтал келеді.
Осылардың ішіндегі маңыздысы цитотоксикалық Т-лимфоциттердің индукциясы (Т-киллер). Олар вирустық антиегндерді клетка антигенінің комплексінің арасынан ажыратылады және зақымдалған клетканы, вирусты бөлшектеп бірге ыдыратады.
Спецификалық емес вирустардан қорғаушы факторға интерферондардың индукциясы жатады. Интерферондар – макроорганизм клеткаларында түзілетін белоктар. Интерферондардың 3 класы бар: лейкоцитарлы – альфа-интерферон, фибробластта түзілетін – бета-интерферон, және интерферондар лейкоциттерде түзілетін – гамма-интерферон. Интерферондардың синтезінің индукциясы әртүрлі факторлардың – интерферондардың - әсеріне байланысты. Олардың біріншісі болып вирусты инфекция, әсіресе РНҚ құрамды вирустар жатады. Интерферон клеткалық рибосомалардағы вирус спецификалық белоктардың трансляциясына әсер етеді, оларды тежейді. Интерферонның талғамдылығы вирустың иесіне байланысты. Интерферон клетка бетіндегі рецепторлармен талғамды әрекеттеседі. Мысалы: тышқанның интерфероны адам ағзасы үшін оншалықты пайдалы емес. Ал кейде интерферон басқаша әрекеттесуі мүмкін. Мысалы: адам интерфероны өгіздің клеткасын, өгіз интерферонына қарағанда жақсы қорғайды. Интерферон спецификалық емес, ол вирустардың көп түрінде тиімді қолданылады. Вирустарға қарсы спецификалық емес факторлардың арасында макрофагтардың және комплементтің рольдері маңызды. Макрофагтар вирус спецификалық антиегндерді Т-лимфоциттерге дайындайды, вируспен зақымдалған клетканы ыдыратуға қатысады. Альтернативтік және классикалық жолымен комплемент жүйесі антиденелермен байланысқан вируспен зақымдалған клеткалардың комплексін ыдыратады. Комплементтің спецификалық антиденелері жоқ болса да кейбір күрделі құрылысы вирустарды ыдырататындығы белгілі.
Вирустық инфекцияны бастан кешіргеннен кейін күшті және ұзаққа созылатын иммунитет пайда болады (қызылша, полимиелит, эпидемиялық паротит, шешек), бірақ ол тұрақсыз немесе қысқа мерзімді болып келеді (тұмау, синциальдық вирусты инфекция) жәневирустардың персистенциясы болуы мүмкін (адено-, ұшық вирустық инфекциялары). Иммунокомпетентті клеткаларда кейбір вирустар көбейеді және персистенцияланады. Мысалы: қызылша вирусы макрофагтарда, ЖИТС вирусы Т-хелперде.
ВИРУСТЫҚ ИНФЕКЦИЯЛАРДЫҢ ЛАБОРАТОРЛЫҚ
ДИАГНОСТИКАСЫНЫҢ ПРИНЦИПТЕРІ
Зерттеу әдістері.
1. Вирусологиялық әдіс – патологиялық материалдағы вирусты анықтау.
- Серологиялық әдіс – аурудың қан сарысуынан вирусқа қарсы антиденелерді анықтау.
- Вирустық инфекциялардың экспресс-диагностикасы: жылдамдатылған серологиялық және вирусоскопиялық әдіс.
| Әдістер | Зерттеу мақсаты |
| Вирусологиялық әдіс | Вирусты биологиялық сезімтал обьектілерден НР, РТГА, РТГ, ИФА, РИФ, РИА, РСК және басқа да реакциялар арқылы анықтайды. |
| Серодиагностика | Вирустарға қарсы қан сарысуындағы антиденелер титрін НР, РТГА, РИФ, ИФА, РИА, РСК реакциялары арқылы анықтайды. |
| Экспресс-диагностика | Клиникалық материалда вирус бөлшектерін немесе вирус спецификалық антигендерді РИФ, ИФА, РИА, ЭМ, ИЭМ және басқа да реакциялар арқылы анықтайды. |
Вирусологиялық зерттеу әдісі 2 кезеңнен тұрады:
А) клиникалық материалдан вирустарды бөліп шығару
Б) бөлінген вирустардың түрлерін анықтау немесе салыстыру.
Вирусты анықтау үшін алынатын зерттеу материалы әртүрлі болады, ол инфекциялық процестің динамикасына және вирустың тропизміне байланысты. Мысалы: тұмау немесе ұшық болса жұтқыншақтан бөлінетін шырыш, везикуладан қырып алынған материал, жұлын сұйығы, ЖИТС инфекциясында Т-лимфоциттердің субпопуляциясы, қан сарысуы, сперма; құтыру ауруында зақымдалған адамның сілекейі және өлгендердің ми тіндері, сілекейдегі бөлінділері зерттеледі.
Вирусты анықтау мақсатында патологиялық материалмен сезімтал лабораториялық жануарларды, тауық эмбриондарын және торша өсіндісіне жұқтырады: Мысалы: тұмау инфекциясында вирус бар материалды торша өсіндісіне егіп немесе күзендер мен тышқандардың мұрын қуысы арқылы жұқтыруға болады. Ол тауық эмбрионының аллантоис немесе амнион қуыстарына енгізу арқылы жұқтыруы ең тиімдісі. ЖИТС кезінде тек қана торша өсіндісінің яғни Т-хелпер пайдаланылады. Вирус лимфоциттер және макрофагтар сирек қолданылады, бұл торшалардың ішінде вирус репродукциясының активтілігі төмен; ұшық инфекциясы кезінде вирусты өсіру үшін торша өсіндісі немесе лабораториялық жануарлар қолданылады. Үй қоянын және тышқандарды құтыру инфекциясын дәлелдеу үшін де қолданылады.
Вирусологиялық зерттеуде біркелкі жануарларды пайдаланған дұрыс. Өйткені олардың генотиптік және фенотиптік қасиеттері ұқсас инфекцияға өте сезімтал.
Патологиялық материалда вирус болған кезде жануарларда инфекциялық аурулардың клиникалық белгілері мен өзгерістері пайда болады. Мысалы: ұшық инфекциясымен құрсақ ішіне зақымдалған тышқандар 3-4 күннен кейін өледі. Егер материалды үй қоянының немесе теңіз шошқысының көзінің мөлдір қабатына жұқтырса, онда қабыну процесі – кератоконьюктивит пайда болады. Құтыру вирусымен зақымдалған тышқандардың аяқ-қолдары жансызданады да өледі. Тауық эмбрионының қабықтарын, аллантоис, амнион қуыстарында вирустың көбейгенінен эмбрион тұқымы зақымданып одан әрі өсуі тоқтатылады. Зақымдалған эмбрионда мынадай өзгерістер байқалады:
1. ақ дақтар – хорион-аллантоистық қабықтағы төмпешіктердің пішіні мен мөлшері өзгереді (тұмау, шешек вирустары), амнион қабығында патоморфологиялық өзгерістер және эмбрионның өлуі.
2. көзге көрінетін өзгерістер болмаса вирусты анықтау үшін тауық эмбрионының аллантоис немесе амнион қуыстарының сұйығымен гемагглютинация реакциясы –РГА қойылады. Көбінесе ажырасқан аллантоис қуысындағы сұйықтарға бірдей көлемде 1% тауық эритроциттері қосылады. Аллантоис қуысындағы сұйықтағы вирустар әсерімен байланысты эритроциттердің агглютинациясы микропланшеттер лункасының түбінде «қол шатыр (зонтик)» тәрізді анықталады. Агглютинация жоқ кезде – эритроциттер лунка түбінде – «түйме» тәрізді орналасады.
Торша өсіндісіндегі патоморфологиялық өзгерістері мынадай: вирустар репродукциясы ядроның немесе цитоплазманың құрылымдарын зақымдап өзгертеді немесе торшаны жансыздандырады. Сондықтан, микроскоп арқылы вирус зақымданған торшада мына өзгерістерді анықтауға болады:
1. Торша ішіне тән патоморфологиялық өзгерістер. Мысалға, құтыру ауруында нейрондардың цитоплазмасында Бабеш-Негри денешіктері анықталады, шешек ауруында – көздің мөлдір қабығының эпителиальдық торшаларының цитоплазмасында – Гварниери денелері, аденовирустық және ұшық инфекцияларында зақымдалған торшалардың ядросында қосындылар анықталады. Бұл қосындылар вирус бөліктері жинақталуы немесе оның компоненттерінің торша материалдарымен жиналуы;
2. вирустың цитопатиялық әсерінен торшаның ісінуі, солуы немесе торшаның дегенеративті өзгеруі қабығының деструкциясымен байқалады. Өлген торшалар адгезивтік қасиетінен айырылып шынының бетінен түсіп қалады. Сондықтан бір қабатты торша өсіндісінде біркелкі торшалардың арасында «алаңдар» немесе бос орындар көрінеді. Симпластар (синцитий) – көп ядролық аз өмір сүретін торшалар вирустардың цитопатиялық әсерінің ерекше белгісі деп саналады, олар ЖИТС, қызылша инфекцияларына тән.
3. зақымдалған торша ішіндегі вирустар гемадсорбция – Ргадс реакциясы арқылы анықталады. Вирустық компоненттер, оның ішінде эритроциттерді агглютинациялайтын гемагглютининдер зақымдалған торшаның мембранасына бекінеді. Сондықтан вирустармен зақымдалған торшалық суспензияға қосылған эритроциттердің 1% қоспасы торша мембранасына адсорбцияланады. Мұндай құбылыс тұмау, қызылша, шешек, аденовирустық және басқа инфекцияларда байқалады. Вирустармен зақымдалған торшалардың гемадсорбциялық қасиеті цитопатиялық өзгерістерден бұрын пайда болады, сондықтангемадсорбция реакциясын вирусты ерте анықтау үшін пайдалануға болады:
4. бляшкалар әдісі, бұл вирустың цитопатиялық әсерін байқау модификациясы. Вирус әсерімен байланысты бір қабатты торша өсіндісінің дөңгелек сияқты бүліншегін бляшкалар деп атайды. Бұл әдісте бір қабатты торшаларды вирустардың аз концентрациясымен зақымдайды және торша бетіндегі вирустық бөлшектерін агармен ұстатады (фиксация), ал агарға витальді бояғыш – нейтральды қызыл қосылған. Бұл жағдайда вирустардың цитопатиялық әсері бір жерге топталған, ал өлген торшалар бояуды ұстау қасиетін жоғалтып түссізденеді. Соның әсерінен бір қабатты торшалардың мөлдір емес қызғылт фонында айқын бляшкалар пайда болады. Олар мөлдір, түссіз дөңгелек таңба сияқты болады. Полимиелит, шешек, қызылша және басқа да инфекцияларды анықтау үшін осы әдіс қолданылады:
5. түрлі-түсті сынама тіндік торшаларды өсіру үшін қолданылады Игла, 199 орталар құрамында қоректік заттармен қатар индикатор – фенолрот болады, егер рН-7,4-7,6 дәрежесінде болса қызыл түсті береді. Қоректік ортада ұлғайып көбейген торшалар зат алмасуы нәтижесінде қышқыл заттарды бөліп шығарады. Бөл себептен қоректік ортаның рН 6,9-7,0 дейін төмендегеннен кейін индикатордың яғни қоректік ортаның түсі қызылдан сарыға өзгереді. Егер қоректік ортадағы торша өсіндісі және вирус – құрайтын материал бір мезгілде кіргізілсе вирустар торшаны зақымдап, дегенеративтік өзгеріс туғызады немесе торшалар жансызданады. Сондықтан метоболиттік процестер тоқталып рН өзгермейді және орта түсі қызыл болып қалады, яғни түрлі-түсті сынама нәтижелі болды деп саналады.
САЛЫСТЫРУ (ИДЕНТИФИКАЦИЯ)
Вирус түрін салыстырып анықтау жолы лаборатория жануарларында нейтралдау реакциясы арқылы жүреді, бірақ бұл әдіс сирек қолданылады. Бұл үшін экспериментальді жануар ағзасына зерттелетін вирус және вирус нейтралдайтын антиденелер бар түрлі спецификалық сарысу қоспасы әртүрлі әдісімен енгізеді. Егер жануарлар ауырмаса, онда вирустардың антиденелермен нейтралдауын байқатады. Белгілі спецификалық антиденелер арқылы вирустың типі анықталады.
Вирустың нозологиялық тәндігі, тауық эмбрионында бөлінген және РГА реакциясында білінген гемагглютинацияны тежеу реакциясымен (РГТА) анықталады. Гемагглютининдердің эритроциттерді агглютинациялау қасиетіне РГА реакциясы негізделген. Бірақ вирусты, құрамында вирусқа бекітілген белоктарды нейтралдайтын антиденелер бар иммунды сарысуымен алдын-ала қосып, кейін эритроциттерді қосса гемагглютинация реакциясы байқалмайды – эритроциттер «түйме» тәрізденіп шұңқыр түбіне тұнады. Бұл жағдай гемагглютинацияны тежеу реакциясын (РТГА) нәтижелі деп саналады. РГА – бұл физика-химиялық реакция, ал РТГА – иммунологиялық реакцияның бір түрі – нейтралдау реакциясы. РТГА реакциясын тек қана вирустың түрін анықтағанда емес, сондай-ақ вирус спецификалық антиденелерді және олардың титрін қан сарысуында айқындауға қолданылады.
Нейтралдау реакциясы торша өсіндісіндегі вирустың идентификациясын өткізгенде қолданылады:
1. нейтралдау реакциясында түрлі-түсті сынама қолданылады. Түрлі-түсті сынаманы өткізу үшін белгілі компонент – торша өсіндісіне (Игла немесе 199 қоректік орта) және вирусты материал түріне спецификалық сарысу қолданылады. Вирустың бөліктері антиденелермен нейтралданса бұл реакция нәтижелі деп саналады. Тіндік торшалар вирустармен зақымданбай өмір сүруге бейім қалпында қалса, олардың зат алмасуынан қышқыл заттар пайда болады. Сондықтан қоректік ортада рН төмендейді және оның қызыл түсі сарыға ауысады:
2. цитопатиялық әсерге байланысты нейтралдау реакциясы. Реакция компоненттері: бір қабатты торша өсіндісі, вирус құрайтын материал және белгілі түрге спецификалық сарысу. Вирустық антигендер антиденелермен нейтрализациялағанда тіндік торшалары бұзылмай қалады. Сондықтан, микроскопта цитопатиялық өзгеріс жоқ, мұндай реакция нәтижелі деп саналады.
3. вирустарда «бляшка» әдісімен үқсастыру және оларды анықтау. Бұл әдіс цитопатиялық әсерлі нейтрализация реакциясының модификациясы болып табылады. Реакция нәтижелі болғанда – дегенеративтік өзгерістер – бляшкалар бір қабатты торшаларда болмайды:
4. гемадсорбцияны тежеу реакциясында вирус нейтралдау антиденелер арқылы вирус түрлерін анықауға болады. Егер бір қабатты торшаларға зерттелетін вирус құрайтын материалды және белгілі түрге спецификалық сарысу енгізгеннен кейін қосылған эритроциттер тіндік торшалар бетіне адсорбцияланбаса, онда реакция нәтижелі деп саналады. Себебі, антиденелер вирустық бөліктерді нейтралдайды, тіндік торшалар зақымдалмайды және олардың сыртқы мембранасында вирустық гемагглютининдер жоқ. Сондықтан эритроциттер тіндік торшалардың мембранасында жиналмайды, гемадсорбция тежеледі. Теріс реакция кезінде антигендермен антиденелердің сәйкестігі болмағанда вирустар тіндік өсіндіні зақымдайды, торшалы мембранаға гемагглютининдер бекітіліп эритроциттер торша маңына жиналады.
Серологиялық зерттеу кезінде антиденелердің титрін анықтау үшін сырқаттың қан сарысуы екі рет 7-10 күн аралығында қайталап тексереді. Қайталап қан сарысуын зерттеудің қажеттілігі мынау: Вирусқа қарсы антиденелердің титрі бірінші рет зерттелген қанда вакцинация немесе ертеде болған жүқпалы аурумен байланысты болуы мүмкін. Егер инфекция динамикасында антиденелердің титрі жоғарыласа ол айтарлықтай дәлел. Антиденелердің титрін 4 есе жоғарылағаны вирустық инфекцияны дәлелдейді.
Серологиялық зерттеу жабдықтары: 1. зерттелетін қан сарысуы, яғни онда вирус нейтралдайтын антиденелерді анықтау; 2. белгілі вирус спецификалық антигендер немесе вирус бөліктері; 3. бұл биологиялық обьект одан жоғарыда көрсеткен зерттелетін қан сарысуының және белгілі вирус бөліктерінің өзара қатынасуын, яғни нейтрализация реакциясының нәтижесін көреміз. Обьект ретінде лабораториялық жануарлар, тауық эмбриондары және торша өсінділері болуы мүмкін. Оң реакция кезінде түрге спецификалық антиденелер, зерттелетін қан сарысуындағы вирустарды нейтралдайды, сондықтан:
1. лабораториялық жануарлар ауырмайды, әрі өлмейді.
2. тауық эмбриондары да патологиялық өзгеріссіз болмайды;
3. бір қабатты торша өсіндісінде ЦПӘ, бляшкалар, гемадсорбция байқалмайды. Торша тіндік өсіндісінде интактілі болып қалады. Түрлі-түсті сынама торша суспензиясының өмір сүру бейімділігін дәлелдейді (қоректік ортаның түсі сары), зерттелген сарысуда табылған антиденелер вирустарды нейтралдайды.
Экспресс-диагностика. Бұл әдіс клиникалық материалды алғаннан кейін бірнеше сағат арасында жауап алуға мүмкіндік береді. Аурулардан алынған материалдан вирусты анықтауға болады. Экспресс әдістің негізі - клиникалық материалды зерттеу. Науқастан алынған материалдағы (қан, шырышты қабықтың жағындысы, үлкен және кіші дәрет) вирустарды сезімтал биологиялық жүйелер арқылы (торша өсіндісі, тайық эмбрионы, лабораториялық жануарлар) өсіріп көбейтудің қажеті жоқ.
1. Электронды-микроскопиялық зерттеу (ЭМ). Бұл әдісті әдетті тәсілдермен өсіруге келмейтін (коронавирустар, ротавирустар, А және В гепатит вирустары) немесе типтік морфологиядағы (шешек вирусы) вирустарды анықтаған кезде ерекше бағалы. ЭМ пайдалануы мынадай жағдайлармен шектеледі, егер клиникалық материалда вирус бөлшектерінің концентрациясы 104-105 мл кем болмау керек. Ұқсас вирустарды ажыратып анықтауға электронды-микроскопиялық әдіс қолданылмайды.
2. Иммунды электронды-микроскопиялық әдіс (ИЭМ). Вирус бөлшектерімен антиденелердің байланысқанында иммунды комплекстер пайда болады. Оларды центрифуга арқылы тұнбаға түсіреді. Электронды-микроскоппен зерттегенде вирустардың жиынтығы көрінеді, ал антиденелер айқын көрініп тұрмаған соң байқалмайды. Белгілі түрге спецификалық антиденелер арқылы иммунды комплекстегі вирустарды анықтауға болады.
3. Жарықты микроскоппен зерттеу. Бұл әдіс басқалар мен салыстырғанда сирек қолданады. Ірі шешек вирусын Морозов әдісімен боялған микропрепаратта көруге болады. Бұл әдіспен Романовскии-Гимзе және басқа әдістермен боялған гиспопрепаратты клетка ішіндегі қосындылар анықталады. Мысалы: зақымдалған нейрондарда Бабеш-Негри денешіктерін осы әдістермен бояғанда байқалады.
4. Иммунофлюоресценция реакциясы – РИФ көптеген вирус инфекцияларының экспресс-диагностикасының бірден бір әдісі. Себебі, люминесцентті микроскоптың кең қолдануы, реакция техникасының қиын еместігі, әдістің жоғары сезімталдығы. РИФ немесе Кунс реакциясының негізін флюоресцентиоционат (ФИТЦ) немесе басқа флюорохромдардың антиденелерімен химиялық комплекс байланысуы, ФИТЦ-пен химиялық байланысқан антиденелер (иммунды сарысулардың құрамында) иммунологиялық ерекшелігін сақтайды және белгілі антигендермен өзара байланысқа түседі. Антигендер мен антиденелердің комплекстерін микропрепаратты сары-жасыл жарық арқылы люминесценттік микроскоппен өте оңай анықтауға болады.
А) иммунофлюоресценцияның түзу реакциясы – зерттелген антигендерге қарсы иммунофлюоресценттік сарысулар пайдаланылады
Б) РИФ түзу емес реакциясы – екі түрлі сарысулар пайдалануға негізделген. Басында зерттелетін антигенге қарсы флюоресценттік емес антиденелер қолданады, ал реакцияның екінші кезеңінде пайда болған антиген – антиденелер комплексін ФИТЦ-пен байланысқан антисары су қосылады. Бұл ФИТЦ-пен байланысқан сарысу (түрге қарсы сарысу) құрамында реакцияның бірінші кезеңінде қолданған флюоресценттік емес антиденелерге немесе гамма-глобулиндерге қарсы анти-антиденелер болады.
Егер, РИФ бірінші кезеңінде қолданған антисары су қойлардан иммунизация арқылы алынса, екінші кезеңінде қолданатын түрге қарсы сарысу дайындау үшін қойдың гаммаглобулиндерімен басқа түрді ажыратады (үй қояны, есек және т.б.) иммунизациялайды. Түрге қарсы сарысу құрамындағы антиденелер ФИТЦ-пен химиялық байланысқан. Сонымен зерттелген антигендерді екінші қабатпен антиглобулиндер жабады (бірінші қабат - флюоресценттік емес антиденелер, олар антиглобулиндері бар сарысуға антиген ретінде болады). Реакция нәтижесінде люминесценттік микроскопта антиген-антидене-анти-антиденелер комплекстері көрінеді. Егер иммунофлюоресценцияның түзу реакциясында әрбір вирус антигендеріне қарсы дереу антисарысуларды ФИТЦ-пен химиялық байланыстыру керек болса, ал түзу емес реакциясында флюорохром тек түрге қарсы сарысумен байланысқан.
РИФ техникасы:
А) реакцияның түзу варианты. Мысалға, респираторлы вирус инфекциясында вируспен зақымдалған торшаларды назофарингиальді секреттен алуға болады. Назофарингиальді секреттегі торшаларды сұйықтан бөліп алып фосфатты буферлі ерітіндімен шаяды. Торша тұнбасына фосфатты буферлі ерітіндісінің бірнеше тамшысы қосылады содан 3-4 жұғын-препаратты дайындайды. Жұғындыларды ауада кептіреді әрі ацетонмен бекітеді. ФИТЦ-пен бекітілген құрамында вирусқа қарсы антиденелер бар сарысудың жұмысты ерітіндісімен препаратты бояйды. Термостатта 30мин. экспозициядан кейін препаратты 3 рет фуцермен, содан соң бір рет дистилденген сумен шаяды. Препарат люминесцентті микроскоппен зерттеледі. Оң реакцияда препарат сары-жасылмен жарқырайды, теріс реакцияда жарқырамайды.
Б) түзу емес реакция варианты. Дайындалған жұңынға құрамында вирустарға қарсы антиденелері бар сарысу тамызады, 30 мин соң экспозиция, ФБР-мен және дистилденген сумен шаяды, оның үстіне ФИТЦ-пен байланысқан антисарысу тамызады.
5. Иммуноферментті анализ – ИФА, антигендер мен спецификалық антиденелердің қатынасына негізделген. Реакцияда ферменттермен белгіленген вирустарға қарсы немесе түрге қарсы антиденелер пайдаланылады. Антигендер және ферменттер байланысқан антиденелер комплекстері спецификалық субстрат (хромоген) арқылы анықталады. Түссіз хромоген ферментпен байланысып бояулы жағдайда өтеді.
Антигендер+ антиденелер (фермент)+ хромоген= бояу.
Антигендер+ антиденелер комплекстерінің саны материалдың боялуының дәрежесіне тура пропорциональді.
Реакцияның бірінші компоненті – антиген, қатты фазаға тұнады. Полистирольды пластинкалардың тегіс түбін қатты фаза деп атайды. Арнайы буфер ерітінділерді инкубациялағанда антиген электростатикалық күш әсерімен қатты фазаға адсорбцияланады.
Пероксидаза, сілтілі фосфатаза, цитохром С және басқа ферменттер антиденелерді белгілеу үшін пайдаланылады. Ферменттер байланыстын субстрат ретінде хромогендер – ортофенилендиамин, ортотолуидин, бензидин және басқа түрлері қолданылады.
ИФА әдістерінің ішінде түзу және түзу еме
|
|
|
|
|
Дата добавления: 2014-10-15; Просмотров: 2653; Нарушение авторских прав?; Мы поможем в написании вашей работы!