КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Ионные каналы клеточных мембран
|
|
|
|
МЕХАНИЗМЫ ГЕНЕРАЦИИ ПОТЕНЦИАЛА ДЕЙСТВИЯ
Экспериментальные данные по генерации биопотенциалов и общие принципы возбудимости биологических мембран, изложенные в главе 3, получили математическое описание, биофизическое обоснование и структурную интерпретацию для ряда важных объектов: возбудимых мембран нервных волокон и мембран клеток сердечной мышцы - кардиомиоцитов.
В данной главе рассмотрены математические модели, раскрывающие механизмы генерации биопотенциалов, а также структура и свойства элементарного проводника в биомембране - ионного канала.
Модель возбудимой мембраны по теории Ходжкина-Хаксли предполагает регулируемый перенос ионов через мембрану. Однако непосредственный переход иона через липидный бислой весьма затруднен, а следовательно, был бы мал и поток ионов.
Это и ряд других соображений дали основание считать, что в мембране должны быть некоторые специальные структуры — проводящие ионы. Такие структуры были найдены и названы ионными каналами. Подобные каналы выделены из различных объектов: плазматической мембраны клеток, постсинаптической мембраны мышечных клеток и других объектов. Известны также ионные каналы, образованные антибиотиками.
Основные свойства ионных каналов:
1) селективность;
2) независимость работы отдельных каналов;
3) дискретный характер проводимости;
4) зависимость параметров каналов от мембранного потенциала.
Рассмотрим их по порядку.
1. Селективностью называют способность ионных каналов избирательно пропускать ионы какого-либо одного типа.
Еще в первых опытах на аксоне кальмара было обнаружено, что ионы Na+ и Кт по-разному влияют на мембранный потенциал. Ионы К+ меняют потенциал покоя, а ионы Na+ - потенциал действия. В модели Ходжкина-Хаксли это описывается путем введения независимых калиевых и натриевых ионных каналов. Предполагалось, что первые пропускают только ионы К+, а вторые - только ионы Na+.
Измерения показали, что ионные каналы обладают абсолютной селективностью по отношению к катионам (катион-селективные каналы) либо к анионам (анион-селективные каналы). В то же время через катион-селективные каналы способны проходить различные катионы различных химических элементов, но проводимость мембраны для неосновного иона, а значит, и ток через нее, будет существенно ниже, например, для Na+-кaнала калиевый ток через него будет в 20 раз меньше. Способность ионного канала пропускать различные ионы называется относительной селективностью и характеризуется рядом селективности - соотношением проводимостей канала для разных ионов, взятых при одной концентрации. При этом для основного иона селективность принимают за 1. Например, для Na+-канала этот ряд имеет вид:
Na+: К+ = 1: 0,05.
2. Независимость работы отдельных каналов. Прохождение тока через отдельный ионный канал не зависит от того, идет ли ток через другие каналы. Например, К+-каналы могут быть включены или выключены, но ток через Nа+-каналы не меняется. Влияние каналов друг на друга происходит опосредованно: изменение проницаемостей каких-либо каналов (например, натриевых) меняет мембранный потенциал, а уже он влияет на проводимости прочих ионных каналов.
3. Дискретный характер проводимости ионных каналов. Ионные каналы представляют собой субъединичный комплекс белков, пронизывающий мембрану. В центре его существует трубка, сквозь которую могут проходить ионы. Количество ионных каналов на 1 мкм2 поверхности мембраны определяли с помощью радиоактивно меченного блокатора натриевых каналов - тетродотоксина. Известно, что одна молекула ТТХ связывается только с одним каналом. Тогда измерение радиоактивности образца с известной площадью позволило показать, что на 1 мкм2 аксона кальмара находится около 500 натриевых каналов.
Те трансмембранные токи, которые измеряют в обычных экспериментах, например, на аксоне кальмара длиной 1 см и диаметром 1 мм, то есть площадью 3*107 мкм2, обусловлены суммарным ответом (изменением проводимости) 500•3•107-1010 ионных каналов. Для такого ответа характерно плавное во времени изменение проводимости. Ответ одиночного ионного канала меняется во времени принципиально иным образом: дискретно и для Nа+-каналов, и для К+-, и для Са2+-каналов.
Впервые это было обнаружено в 1962 г. в исследованиях проводимости бислойных липидных мембран (БЛМ) при добавлении в раствор, омывающий мембрану, микроколичеств некоторого вещества, индуцировавшего возбуждение. На БЛМ подавали постоянное напряжение и регистрировали ток I(t). Запись тока во времени имела вид скачков между двумя проводящими состояниями.
Одним из эффективных методов экспериментального исследования ионных каналов стал разработанный в 80-е годы метод локальной фиксации потенциала мембраны ("Patch Clamp"), (рис. 10).
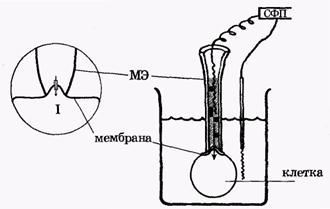
Рис. 10. Метод локальной фиксации потенциала мембраны. МЭ — микроэлектрод, ИК - ионный канал, М — мембрана клетки, СФП - схема фиксации потенциала, I - ток одиночного канала
Суть метода заключается в том, что микроэлектрод МЭ (рис. 10) тонким концом, имеющим диаметр 0,5-1 мкм, присасывается к мембране таким образом, чтобы в его внутренний диаметр попал ионный канал. Тогда, используя схему фиксации потенциала, можно измерять токи, которые проходят только через одиночный канал мембраны, а не через все каналы одновременно, как это происходит при использовании стандартного метода фиксации потенциала.
Результаты экспериментов, выполненных на различных ионных каналах, показали, что проводимость ионного канала дискретна и он может находиться в двух состояниях: открытом или закрытом. Переходы между состояниями происходят в случайные моменты времени и подчиняются статистическим закономерностям. Нельзя сказать, что данный ионный канал откроется именно в этот момент времени. Можно лишь сделать утверждение о вероятности открывания канала в определенном интервале времени.
4. Зависимость параметров канала от мембранного потенциала. Ионные каналы нервных волокон чувствительны к мембранному потенциалу, например натриевый и калиевый каналы аксона кальмара. Это проявляется в том, что после начала деполяризации мембраны соответствующие токи начинают изменяться с той или иной кинетикой. Этот процесс происходит следующим образом: Ион-селективный канал имеет сенсор — некоторый элемент своей конструкции, чувствительный к действию электрического поля (рис. 11). При изменении мембранного потенциала меняется величина действующей на него силы, в результате эта часть ионного канала перемещается и меняет вероятность открывания или закрывания ворот — своеобразных заслонок, действующих по закону "все или ничего". Экспериментально показано, что под действием деполяризации мембраны увеличивается вероятность перехода натриевого канала в проводящее состояние. Скачок напряжения на мембране, создаваемый при измерениях методом фиксации потенциала, приводит к тому, что большое число каналов открывается. Через них проходит больше зарядов, а значит, в среднем, протекает больший ток. Существенно, что процесс роста проводимости канала определяется увеличением вероятности перехода канала в открытое состояние, а не увеличением диаметра открытого канала. Таково современное представление о механизме прохождения тока через одиночный канал.
Плавные кинетические кривые токов, регистрируемых при электрических измерениях на больших мембранах, получаются вследствие суммации многих скачкообразных токов, протекающих через отдельные каналы. Их суммирование, как показано выше, резко уменьшает флуктуации и дает достаточно гладкие зависимости трансмембранного тока от времени.
Ионные каналы могут быть чувствительны и к другим физическим воздействиям: механическим деформациям, связыванию химических веществ и т.д. В этом случае они являются структурной основой, соответственно, механорецепторов, хемо-рецепторов и т.д.
Изучение ионных каналов в мембранах есть одна из важных задач современной биофизики.
|
|
|
|
|
Дата добавления: 2014-10-23; Просмотров: 1313; Нарушение авторских прав?; Мы поможем в написании вашей работы!