КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Химия нуклеиновых кислот
|
|
|
|
Форма белковых молекул. Данные различных видов анализа указывают на существование в природе глобулярных (шарообразных) и фибриллярных (нитевидных) белков. В настоящее время общие представления о форме белковых молекул в основном подтвердились, однако только современные методы исследования позволили установить детали пространственной конфигурации (трехмерной структуры) белковых молекул. Благодаря применению сканирующей микроскопии и рентгеноструктурного анализа удалось в деталях расшифровать не только полную пространственную структуру, форму, но и степень асимметрии белковых молекул во всех трех измерениях. Оказалось, что даже глобулярные белки крови (гемоглобин, альбумины и глобулины) являются асимметричными в указанных измерениях.
Молекулярная масса белков. Белки относятся к высокомолекулярным соединениям, в состав которых входят сотни и даже тысячи аминокислотных остатков, объединенных в макромолекулярную структуру. Молекулярная масса белков колеблется от 6000 (нижний предел) до 1000000 и выше в зависимости от количества отдельных полипептидных цепей в составе единой молекулярной структуры белка. Такие полипептидные цепи получили название субъединиц.
Аминокислотный состав и последовательность аминокислот выяснена для многих тысяч белков. В связи с этим стало возможным вычисление их молекулярной массы химическим путем с высокой точностью. Однако для огромного количества встречающихся в природе белков химическое строение не выяснено, поэтому основными методами определения молекулярной массы все еще остаются физико-химические методы (гравиметрические, осмометрические, вискозиметрические, электрофоретические, оптические и др.). На практике наиболее часто используются методы седиментационного анализа, где определение молекулярной массы белков проводят в ультрацентрифугах и вычисляют её по скорости седиментации молекул белка или седиментационному равновесию.
Денатурация белков. Природные белковые тела наделеныопределенной, строго заданной пространственной конфигурацией и обладают рядом характерных физико-химических и биологических свойств при физиологических значениях температуры и рН среды.Под влиянием различных физических и химических факторов белки подвергаются свертыванию и выпадают в осадок, теряя нативные свойства.
Таким образом, под денатурацией следует понимать нарушение общего плана уникальной структуры нативной молекулы белка, преимущественно ее третичной структуры, приводящее к потере характерных для нее свойств (растворимость, электрофоретическая подвижность, биологическая активность и т.д.). Большинство белков денатурирует при нагревании их растворов выше 50–60°С (рис.6).
Внешние проявления денатурации сводятся к потере растворимости, особенно в изоэлектрической точке, повышению вязкости белковых растворов, увеличению количества свободных функциональных SH-групп и изменению характера рассеивания рентгеновских лучей. Наиболее характерным признаком денатурации является резкое снижение или полная потеря белком его биологической активности (каталитической, антигенной или гормональной). При денатурации белка, разрушаются в основном нековалентные связи (в частности, гидрофобные взаимодействия и водородные связи).
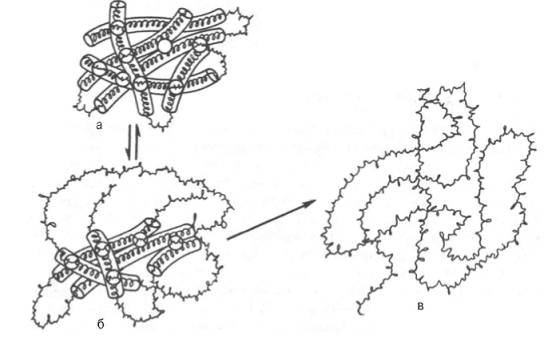
а - исходное состояние; б - начинающееся обратимое нарушение молекулярной структуры; в - необратимое развертывание полипептидной цепи.
Рисунок 6 -Денатурация белковой молекулы (схема)
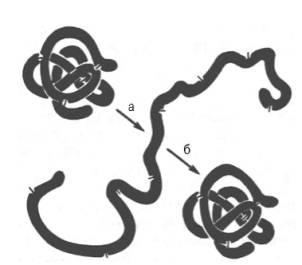
а - развертывание (мочевина + меркаптоэтанол); б - повторное свертывание
Рисунок 7- Денатурация и ренатурация рибонуклеазы (по Анфинсену)
При непродолжительном действии и быстром удалении денатурирующих агентов возможна ренатурация белка с полным восстановлением исходной трехмерной структуры и нативных свойств его молекулы (рис. 7), включая биологическую активность. Таким образом, при денатурации белковая молекула полностью теряет биологические свойства, демонстрируя тем самым тесную связь между структурой и функцией. Для практических целей иногда используют процесс денатурации в «мягких» условиях, например при получении ферментов или других биологически активных белковых препаратов в условиях низких температур в присутствии солей и при соответствующем значении рН. При лиофилизации белков (высушивание в вакууме путем возгонки влаги из замороженного состояния) для предотвращения денатурации часто пользуются химическими веществами (простые сахара, глицерин, органические анионы).
Изоэлектрическая и изоионная точки белков. В изоэлектрической точке суммарный заряд белков, обладающих амфотерными свойствами, равен нулю и белки не перемещаются в электрическом поле. Зная аминокислотный состав белка, можно приближенно определить изоэлектрическую точку (p I); p I является характерной константой белков. Изоэлектрическая точка большинства белков животных тканей лежит в пределах от 5,5 до 7,0, что свидетельствует о частичном преобладании кислых аминокислот. Однако в природе имеются белки, у которых значения изоэлектрических точек лежат в крайних значениях рН среды. В частности, величина р I пепсина (фермент желудочного сока) равна 1, а сальмина (основной белок из молоки семги) – почти 12.
В изоэлектрической точке белки наименее устойчивы в растворе и легко выпадают в осадок. Изоэлектрическая точка белка в сильной степени зависит от присутствия в растворе ионов солей; в то же время на ее величину не влияет концентрация белка.
Раствор белка называется изоионным, если он не содержит никаких других ионов, кроме ионизированных остатков аминокислот белковой молекулы и ионов, образующихся при диссоциации воды. Для освобождения белка от посторонних ионов обычно его раствор пропускают через колонку, наполненную смесью анионо- и катионообменников. Изоионной точкой данного белка принято называть значение рН изоионного раствора этого белка:
[Н]+ + [Р] Z = [ОН]-
где [ Р ] – молярная концентрация белка; Z – средний заряд молекулы. Согласно этому уравнению, изоионная точка белка зависит от его концентрации. Очевидно, поэтому белок, за исключением случая, когда рI равно 7, не может быть одновременно изоэлектрическим и изоионным.
Кислотно-основные свойства. Белки, как и аминокислоты, являются полиамфолитами, проявляя кислотные свойства за счет неионизированных группы – СООН, аммонийных групп – NН3+, тиольных групп – SН. Основные свойства белки проявляют за счет групп –СОО-, аминогрупп –NН2 и др. В водных растворах в зависимости от рН среды белки могут находиться при рН=р I белка в молекулярной, т.е. нейтральной форме, при рН < р I белок проявляет катионные свойства, а при рН > р I проявляется анионная форма.
NН3+ – Рrot – СОО- ↔ NН3+ – Рrot – СОО- ↔ NН3+ – Рrot – СОО-
|+ | |
RН2 RН R-
рН < р I рН = р I рН > р I
катион белка молекула белка анион белка
В зависимости от аминокислотного состава белки подразделяются на «нейтральные» (р I = 5,0-7,0), «кислотные» (р I < 4,0) с повышенным содержанием аспарагиновой и глутаминовой кислоты, и «основные» с повышенным содержанием аргинина, лизина или гистидина (р I >7,5). На основе белков в организме действуют буферные свойства белков.
Буферные свойства белков обусловлены наличием в составляющих их аминокислотах (карбоксикислотах) аминогруппы (NH2-группы). Благодаря ней аминокислоты могут реагировать не только как слабые кислоты, но и как основания, то есть сами проявлять буферные свойства, присоединяя или отдавая ион водорода. Отщепляемый от карбоксильной группы протон может присоединиться к аминогруппе. В результате – молекула аминокислоты принимает дипольную форму (или форму цвиттер-иона), заряжаясь с одной стороны отрицательно, а с другой – положительно, но оставаясь в целом нейтральной. Именно в этой форме аминокислота и проявляет свои буферные свойства. При повышении концентрации протонов в среде (снижение рН) они фиксируются карбоксильной группой, а молекула оказывается положительно заряженной. Наоборот, при падении концентрации протонов третий протон с положительно заряженной стороны молекулы отдается, а вся молекула заряжается отрицательно. Аминокислота диссоциирует с образованием протона и диссоциированной карбоксильной группы.
NН2–R–СООН ↔ NН2–R–СОО- + Н+
Или аминогруппа принимает свободный протон и приобретает форму цвиттер-иона. В избытке протонов молекула заряжается положительно:
Н+ + NH2–R–СOО– ↔ NH3+ –R – СOО–
При дефиците протонов - молекула приобретает отрицательный заряд:
NН3+–R–СОО– ↔ Н+ + NН2–R–СОО–
Буферные свойства белков проявляются в связывании не только протонов, но и других заряженных частиц. Основная масса поступающих в кровоток веществ (красители, жирные кислоты, липиды, водорастворимые наркотики, релаксанты) связывается с белками, проявляя конкурентные отношения. Естественно, при этом уменьшается буферная емкость белков в отношении протонов, и высокая концентрация последних затрудняет освобождение и ослабляет действие веществ, образующих положительные заряды.
Белки, содержащие аспарагиновую и глутаминовую кислоту, вступают в реакцию амидирования белков, при которой обезвреживается аммиак:
NН3+ – Рrot – СОО- + NН3 ↔ NН3+ – Рrot – СОО-
|+ | + Н2О
СН2 – СООН СН2 – СОNН2
Окислительно-восстановительные свойства. Белки относительно устойчивы к мягкому окислению, за исключением содержащих аминокислоту цистеин, так как тиольная группа её легко окисляется в дисульфидную группу, причем этот процесс может носить и обратный характер:
окислитель
2 R–SН R–S – S – R +2е + 2 Н+
Восстановленная восстановитель окисленная
форма форма
В результате этих превращений происходит изменение конформации белка и его нативных свойств. Эти превращения лежат в основе химической завивки волос, так как цистеин и цистин входят в состав белка волос кератина. Сначала волосы обрабатывают восстановителем, чтобы разрушить связи –S – S – цистина и превратить в тиольные группы цистеина. Затем волосы укладывают в локоны и обрабатывают окислителем. При этом образуются дисульфидные связи цистина, которые помогают волосам сохранить их новую форму.
В организме белки, содержащие остатки лизина, пролина, фенилаланина и триптофана, подвергаются ферментативному гидроксилированию при участии кислорода и восстановленной формы кофермента:
NН3+ – Рrot – СОО- + О2 + восстановленная ↔ NН3+ – Рrot – СОО-+ Н2О + окисленная
| форма | форма
RН кофермента RОН кофермента
Также для белков характерны и все цветные (качественные) реакции на аминокислоты.
В наше время трудно назвать область естествознания, которую не интересовала бы проблема структуры и функций нуклеиновых кислот. Несмотря на огромный прогресс, достигнутый в последние десятилетия при изучении химического состава и строения нуклеиновых кислот, много проблем предстоит еще решить для выяснения зависимости между структурой и биологической ролью нуклеиновых кислот. Нет сомнения, что именно на этом пути научного поиска исследования нуклеиновых кислот будут сделаны открытия, имеющие огромное значение для биологии, медицины и всей науки о живом. Эпохальное открытие принципа комплементарности нуклеиновых кислот позволило проникнуть в тайны не только тонкой структуры этих биополимеров, но и механизмов синтеза и воспроизведения биологических макромолекул.
Нуклеиновые кислоты выполняют ряд важных биологических функций, не свойственных другим полимерным веществам. В частности, они обеспечивают хранение и передачу наследственной информации и принимают непосредственное участие в механизмах реализации этой информации путем программирования синтеза всех клеточных белков. Структурные компоненты нуклеиновых кислот выполняют, кроме того, функции кофакторов (коэнзим А, уридиндифосфатглюкоза и др.), аллостерических эффекторов, входят в состав коферментов (никотинамидадениндинуклеотид (НАД), флавинадениндинукле-отид (ФАД) и др.), принимая тем самым непосредственное участие в обмене веществ, а также в аккумулировании (накоплении), переносе и трасформации энергии. Они являются предшественниками вторичных посредников – циклических мононуклеотидов (цАМФ и цГМФ), выполняющих важную функцию в передаче внутриклеточных сигналов.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 1109; Нарушение авторских прав?; Мы поможем в написании вашей работы!