КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Факторы некроза опухолей 1 страница
|
|
|
|
Интерлейкины. Факторы некроза опухолей. Колониестимулирующие факторы. Интерфероны. Ростовые факторы. Хемокины. Биологические эффекты цитокинов. Клиническое применение цитокинов (цитокиновая/антицитокиновая терапия).
Интерлейкины
В 1979 г. на Международном симпозиуме комитета экспертов по лимфокинам в Швейцарии были обобщены накопленные к тому времени данные о лимфокинах и монокинах и сформулирована концепция интерлейкинов (ИЛ) – медиаторов межлейкоцитарных взаимодействий. Тогда же получили порядковые номера два первых интерлейкина – ИЛ-1 и ИЛ-2. К настоящему времени описано 37 интерлейкинов. Все они получены в рекомбинантной форме, и известна их первичная структура.
ИНТЕРЛЕЙКИН-1 (кодируется генами 2-й пары хромосом у человека). ИЛ-1 описан в 1972 г. как фактор, способный индуцировать пролиферацию тимоцитов при совместном действии с митогенами в низких концентрациях.
Клонирование генов ИЛ-1 показало, что существует два различных белка с практически идентичной активностью: ИЛ-1α и ИЛ-1β, которые имеют близкую молекулярную массу (15-17 кД), качественно сходны и являются центральными цитокинами врожденного и адаптивного иммунного ответа и воспаления.
Основные продуценты ИЛ-1 – моноциты и макрофаги. ИЛ-1 образуется также В-лимфоцитами, дендритными клетками, нейтрофилами, клетками Лангерганса, глиальными, эндотелиальными и синовиальными клетками, фибробластами, эпителием кожи и тимуса, в культуре – некоторыми клонами Т-лимфоцитов. Условием выработки ИЛ-1 моноцитами и макрофагами является их активация через Толл-подобные и NOD-рецепторы бактериальными, вирусными и иными продуктами (табл. 16).
Ингибиторами синтеза ИЛ-1 являются: простагландин Е2, глюкокортикоиды; факторы, повышающие уровень цАМФ; TGF-β и др.
Спектр клеток-мишеней для ИЛ-1 очень широк: активированные Т- и В-лимфоциты, макрофаги и нейтрофилы, NK-клетки, клетки эндотелия,
гладкомышечные клетки и клетки хряща (хондроциты), базофилы, плазмоциты, кроветворные клетки, кератиноциты, остеокласты, гепатоциты, клетки островков Лангерганса поджелудочной железы (табл. 16). Все они экспрессируют рецепторы, общие для α- и β-форм ИЛ-1. Условием проявления действия ИЛ-1 является погружение комплекса цитокина с рецептором внутрь клетки.
Биологические эффекты ИЛ-1 можно условно разделить на: иммунологические, воспалительные, кроветворные и межсистемные.
Таблица 16.Стимуляторы продукции ИЛ-1
| Группа | Вещества |
| Компоненты клеточ-ной стенки | липополисахариды, пептидогликаны, мурамилди-пептид, белок А стафилококка |
| Стимуляторы фагоци-тоза | частицы кремния, зимозан, латекс |
| Компоненты иммун-ной системы | гетерогенные антигены, иммунные комплексы, агрегированный IgA; IgЕ, рецептор для IgЕ (CD23), компоненты комплемента С3а, С5а |
| Цитокины | ИЛ-1, 2, 6; TNF, интефероны, ГМ-КСФ |
| Химические соедине-ния | форболовые эфиры, полимиксин В, сапонин, декс-трансульфат |
| Физические факторы | УФ-облучение, прилипание к пластику |
| Другие биологически активные соединения | амилоид Р, тромбин, лейкотриены |
ИЛ-1 причастен к запуску начальных событий адаптивного иммунного ответа (рис. 48), в частности, к вовлечению в него Т-хелперов. Среди Т-клеток рецепторы для ИЛ-1 в наибольшей степени экспрессируют Тх2, продуцирующие ИЛ-4 и 5, но не ИЛ-2 и IFN-γ. Однако, ИЛ-1 способствует также экспрессии генов ИЛ-2 и рецепторов для него. По-видимому, первый из этих эффектов реализуется посредством косвенных механизмов. ИЛ-1 участвует и в контактном взаимодействии Т-лимфоцитов с макрофагами. ИЛ-1 наряду с другими цитокинами (ИЛ-4) вызывает пролиферацию активированных В-лимфоцитов, их дифференцировку в плазматические клетки и регуляцию переключения изотипов иммуноглобулинов. Совместно с ИЛ-2 и IFN-γ повышает активность NK-клеток.
ИЛ-1 стимулирует миелопоэз и ранние этапы эритропоэза (поздние – подавляет, будучи антагонистом эритропоэтина). С действием на кроветворение связан и радиозащитный эффект ИЛ-1, проявляющийся при его введении до облучения и усиливающийся при введении через 5 суток после облучения.
ИЛ-1 известен как провоспалительный цитокин. Он способен индуцировать большую часть местных и системных проявлений воспалительной реакции. Это достигается через повышение адгезивности эндотелия сосудов для клеток крови, увеличение прокоагулянтной активности клеток. ИЛ-1 повышает подвижность нейтрофилов; для ряда клеток является хемоаттрактантом; способствует активации клеток в очаге воспаления; усиливает продукцию ими других цитокинов, а также простагландинов; повышает синтез коллагена и фибронектина; стимулирует фагоцитоз и генерацию супероксид-радикалов; вызывает дегрануляцию тучных клеток. Все это способствует развитию экссудативной и пролиферативной составляющих воспалительной реакции.
Таблица 17.Биологическое действие ИЛ-1 на клетки - мишени
| Клетки-мишени | Биологическое действие |
| Зрелые Т-лимфоциты | Индуцирует пролиферацию Тх2 в комбинации со стимуляцией через антигенраспознающий рецептор Т-лимфоцитов |
| Зрелые В-лимфоциты | Индуцирует их дифференцировку в плазматические клетки, пролиферацию и секрецию иммуноглобулинов |
| Предшественники В-лимфоцитов | Индуцирует дифференцировку и экспрессию поверхностного IgМ |
| NK-клетки | Усиление цитотоксичности, повышение экспрессии рецептора к ИЛ-2;стимуляция продукции IFN-γ |
| Макрофаги | Стимуляция фагоцитоза, хемотаксиса, цитотоксичности; генерация супероксиданионов и окиси азота; увеличение экспрессии белков МНС II класса; стимуляция продукции ИЛ-1, ИЛ-6 и TNF-a (аутокринно) |
| Нейтрофилы | Уисление хемотаксиса и дегрануляции, индукция образования активных форм кислорода и окиси азота |
| Фибробласты | Вызывает пролиферацию и индуцирует секрецию коллагена-зы, стромелизина, ИЛ-6 и Г-КСФ, синтез циклооксигеназы и, следовательно, высвобождение простагландинов (подав-ляют экспрессию м-РНК для основных белков) |
| Гладкомышечные клетки | Вызывают пролиферацию, протеолиз мышц; стимуляцию продукции простагландина Е2 |
| Кератиноциты | Вызывают пролиферацию и выделение других цитокинов, например, ИЛ-6 |
| Эндотелиальные клетки | Стимуляция пролиферации, изменение экспрессии поверх-ностных эндотелиальных антигенов; индуцируют высво-бождение TNF и экспрессию/активацию молекул адгезии |
| Остеокласты | Активирует протонную помпу, что приводит к резорбции костей |
| Хондроциты | Индуцирует секрецию металлопротеиназ, что ведет к разрушению хряща и выделению протеингликанов |
| Гепатоциты | Индуцирует секрецию белков острой фазы, снижение синтеза альбумина; увеличение секреции С3-компонента комплемента |
| Гемопоэтические предшественники | Вместе с ГМ-КСФ индуцирует пролиферацию и дифферен-цировку гранулоцитов и моноцитов |
| Базофилы и тучные клетки | Индукция выброса гистамина |
| Клетки поджелудоч-ной железы | Разрушение β-клеток островков Лангерганса |
| Опухолевые клетки | Разрушение клеток |
Обусловливая рассасывание костной и хрящевой ткани, ИЛ-1 участвует в развитии альтернативных проявлений воспалительной реакции. Однако низкие концентрации ИЛ-1 могут способствовать пролиферации остеобластов и кратковременно стимулировать синтез коллагена.
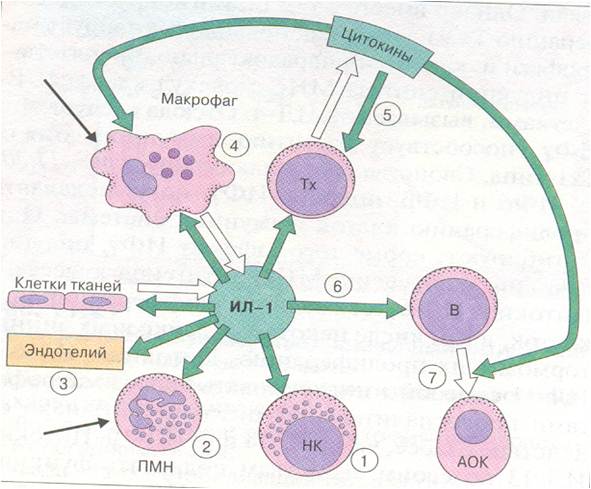
Рисунок 48. Действие ИЛ-1 на клетки иммунной системы
(по А.Ройту и др., 2000г.)
1-повышение активности NK–клеток; 2-повышение метаболической активности полиморфноядерных нейтрофилов (ПМН); 3-индукция синтеза молекул адгезии и прокоагулянтов в эндотелиальных клеток и увеличение проницаемости эндотелия; 4-повышение продукции простагландинов и цитокинов макрофагами; 5-усиление пролиферации Тх, экспрессии рецепторов ИЛ-2 и продукции цитокинов; 6-пролиферация В-лимфоцитов и их дифференцировка в антителообразующие клетки
ИЛ-1 является пирогенным фактором и обусловливает развитие лихорадки. Через действие на миелопоэз ИЛ-1 вызывает нейтрофильный лейкоцитоз. Кроме того, он способствует индукции синтеза белков острой фазы воспаления в гепатоцитах.
Многие эффекты ИЛ-1 осуществляются в кооперации с ИЛ-6 и TNF-α. ИЛ-1 участвует в обмене сигналами между иммунной и нервной, а также нейроэндокринной системами. Он синтезируется в центральной нервной системе клетками глии и сосудистым эндотелием, а кроме того проникает из крови в области III и IV желудочков головного мозга. Рецепторы для ИЛ-1 обнаружены на клетках головного мозга, особенно в извилинах, хориоидных сосудистых сплетениях, гипокампе и гипофизе. ИЛ-1 усиливает экспрессию гена проопиомеланокортина, влияя таким образом на образование ряда гормонов гипоталамуса и гипофиза; стимулирует выработку гормона тимуса тимулина, подавляет образование пролактина, участвует в регуляции сна и поведенческих реакций.
Описано несколько ингибиторов ИЛ-1. Один из них получен из мочи людей с повышенной температурой тела (фебрильный ингибитор ИЛ-1 с молекулярной массой 30 кД). Этот ингибитор взаимодействует с рецептором для ИЛ-1 и отменяет большую часть его эффектов. Другой ингибитор (молек. масса 26 кД) получен из мочи больных моноцитарным лейкозом. Третий мочевой антагонист ИЛ-1 – уромодулин (молек. масса 85 кД) – выделен из мочи беременных женщин.
Наибольший интерес представляет рецепторный антагонист ИЛ-1 (РАИЛ-1) – представитель того же семейства молекул, к которому принадлежат ИЛ-1α и ИЛ-1β. Он имеет молекулярную массу 17 кД и обладает сродством к рецепторам для ИЛ-1. Однако РАИЛ-1 не только не оказывает ИЛ-1-подобного действия, но, наоборот, отменяет большинство его эффектов. В настоящее время он рассматривается как перспективный противовоспалительный фактор локального действия.
Таким образом, ИЛ-1 является цитокином широкого спектра действия и продуцируется преимущественно макрофагами. Он обусловливает пусковые реакции иммунитета, играет ключевую роль в развитии воспаления (участвует в патогенезе большинства острых и хронических воспалительных заболеваний, таких как септических шок, ревматоидный артрит, атеросклероз), участвует в регуляции гемопоэза, является медиатором взаимодействий между иммунной и нервной системами.
ИНТЕРЛЕЙКИН-2 (кодируется генами 4-й пары хромосом у человека). ИЛ-2 описан в 1976 г. как фактор роста Т-клеток. В 1980 г. очищен и охарактеризован биохимически. ИЛ-2 представляет собой гликопротеин с молекулярной массой 15 кД. В молекуле имеется одна дисульфидная связь. У ИЛ-2 меньший спектр биологической активности, чем у ИЛ-1.
На долю CD4+-лимфоцитов приходится 90% клеток-продуцентов ИЛ-2, на долю CD8+ – около 10%. Основными продуцентами ИЛ-2 являются активированные Тх1. Образование ИЛ-2 подавляют глюкокортикоиды, оксимочевина, азатиоприн, ганглиозиды, дезоксиаденозин, а также простагландины и другие факторы, повышающие уровень цАМФ. Лейкотриены и агенты, ингибирующие фосфолипазы и циклооксигеназу, усиливают синтез ИЛ-2. Среди цитокинов способностью повышать продукцию ИЛ-2 обладают ИЛ-1, 6, TNF-α, IFN-γ.
ИЛ-2 обладает относительно узким спектром клеток-мишеней и биологических эффектов. Основными клетками-мишенями для него являются активированные Т- и В-лимфоциты и натуральные киллеры. Главное действие, оказываемое им на лимфоциты, – это митогенный эффект, то есть он обеспечивает размножение лимфоцитов и в некоторой степени стимулирует их дифференцировку: Тх1 (аутокринно), CD8+ цитотоксических Т-лимфоцитов, NK-клеток и В-лимфоцитов, индуцируя синтез IgM и IgG (1, 3).
ИЛ-2 служит дифференцировочным фактором для CD8+ЦТЛ. Это действие ИЛ-2 проявляется позже, чем ростовое, и для его эффективного осуществления требуются дополнительные факторы, такие как ИЛ-6, 7 и ИЛ-12. ИЛ-2 способствует реализации функции Тх1, усиливая выработку IFN-γ, экспрессию протоонкогенов и т.д. Он препятствует развитию иммунологической толерантности и даже отменяет ее.
ИЛ-2 подавляет образование Тх2 и ингибирует синтез продуцируемых ими цитокинов (ИЛ-4, 5 и ИЛ-10).
ИЛ-2 действует как один из ростовых факторов на предварительно активированные В-лимфоциты. Это его влияние усиливают ИЛ-5 и IFN-γ. ИЛ-2 может также повышать синтез IgM, IgG и IgА.
ИЛ-2 увеличивает цитотоксическую активность NK-клеток, расширяя спектр их цитотоксического действия.
ИЛ-2 воздействует также на моноциты, усиливая генерацию активных форм кислорода и оксида азота, а также влияет на процесс кроветворения. Он повышает образование эозинофилов и тромбоцитов, но подавляет миелоидный и эритроидный ростки кроветворения, способствует развитию экстрамедуллярных очагов гемопоэза.
Таким образом, ИЛ-2 в первую очередь представляет собой фактор роста и дифференцировки Т-лимфоцитов и NK-клеток, в меньшей степени В-лимфоцитов, продуцируемый активированными Т-хелперами. Он является важнейшим медиатором адаптивного иммунитета (особенно Т-клеточного) и участвует в реализации иммунной защиты и противоопухолевой резистентности.
ИНТЕРЛЕЙКИН-3 (кодируется у человека генами 5-й пары хромосом, локализованными очень близко от гена, кодирующего гранулоцитарно-макрофагальный колониестимулирующий фактор). Этот цитокин описан в 1981 г. как фактор дифференцировки Т-клеток. Несколько позже было установлено, что ИЛ-3 является фактором, стимулирующим ранние стадии гемопоэза. ИЛ-3 – сильно гликозированный белок с молекулярной массой 21-36 кД. В молекуле имеется одна дисульфидная связь, важная для проявления активности.
Клетками-продуцентами ИЛ-3 являются Тх1 и Тх2, активированные специфическими антигенами, а также ряд других клеток (В-лимфоциты; тучные клетки, активированные перекрестным связыванием цитофильных антител, прикрепленных к их поверхности, со специфическими антигенами; миелоидные клетки; стромальные клетки костного мозга; астроциты головного мозга; кератиноциты).
Клетками-мишенями ИЛ-3 служат в основном юные, в том числе полипотентные, кроветворные предшественники.
ИЛ-3 обеспечивает поддержание пролиферации стволовых клеток и незрелых предшественников лимфоцитов, индукцию дифференцировки макрофагов в костном мозге и стимуляцию дифференцировки Т-лимфоцитов в тимусе.
ИЛ-3 является ростовым фактором для тучных клеток слизистых оболочек и усиливает ростостимулирующую активность ИЛ-4 в отношении тучных клеток соединительной ткани (серозные); усиливает продукцию ими гистамина, вследствие чего ИЛ-3 причастен к развитию аллергических реакций.
ИЛ-3 активирует эозинофилы и тормозит развитие NK-клеток. ИЛ-3 повышает выживаемость клеток за счет блокирования апоптоза.
Таким образом, ИЛ-3 представляет собой продукт в первую очередь активированных Т-лимфоцитов, влияющий главным образом на ранние стадии гемопоэза и ответственный за его экстренную регуляцию. Кроме того, ИЛ-3 также участвует в развитии тучных клеток и подавляет формирование NK-клеток.
ИНТЕРЛЕЙКИН-4 (кодируется у человека генами 5-й пары хромосом, локализованными недалеко от гена для ИЛ-5). ИЛ-4 описан в 1981 г. как фактор, стимулирующий В-лимфоциты. Имеет другие названия: B-cell growth factor I (BCGF-I), B-cell stimulatory factor I (BCSF-I), T-cell growth factor II (TCGF-II), mast cell growth factor II (MCGF-II). ИЛ-4 – это гликопротеин с молекулярной массой 19-22 кД, в его молекуле имеется дисульфидная связь, важная для проявления активности.
Главными продуцентами ИЛ-4 являются Тх2. ИЛ-4 продуцируется также тучными клетками, базофилами и В-клеточными линиями. Кроме того, небольшое количество ИЛ-4 синтезируют Тх0. Глюкокортикоиды в дозах, подавляющих синтез ИЛ-2, повышают образование ИЛ-4. Усиление синтеза ИЛ-4 происходит при старении.
Рецепторы для ИЛ-4 обнаружены на многих клетках. В отношении Т-лимфоцитов ИЛ-4 является как бы двойником-антагонистом ИЛ-2. ИЛ-4 вызывает пролиферацию тимоцитов и активированных Т-клеток, причем ИЛ-4 является основным фактором, обусловливающим дифференцировку CD4+-клеток в направлении Тх2, благодаря чему обеспечивается аутокринная стимуляция пролиферации и дополнительное образование Тх2.
Являясь ростовым фактором для В-лимфоцитов, ИЛ -4 осуществляет переключение синтеза классов иммуноглобулинов на синтез цитофильных IgE и IgG(2,4). Эти эффекты, а также способность ИЛ-4 поддерживать пролиферацию тучных клеток имеют прямое отношение к развитию атопических аллергических реакций. У мышей с мутацией, разрушающей ген ИЛ-4, полностью утрачивается способность к синтезу IgE и цитофильного IgG.
ИЛ-4 повышает экспрессию продуктов МНС II класса и антигенпредставляющую активность вспомогательных клеток. В этом отношении ИЛ-4 является функциональным аналогом IFN-γ, хотя во многих других ситуациях он выступает как его антагонист.
Подавляя функции макрофагов и секрецию ими ИЛ-1, TNF-a и ИЛ-6, ИЛ-4 оказывает противовоспалительное действие. В то же время он повышает цитотоксическую активность макрофагов, способствует миграции в очаг воспаления нейтрофилов, усиливает выработку колоние-стимулирующих факторов. ИЛ-4 стимулирует гемопоэз, способствуя выживаемости кроветворных клеток.
Важным свойством ИЛ-4 является его выраженная противоопухолевая активность in vivo. При местном введении ИЛ-4 индуцирует быструю инфильтрацию опухоли эозинофилами, оказывающими цитотоксическое действие на опухолевые клетки.
ИЛ-4, благодаря индукции ИЛ-5, стимулирует in vivo образование большого количества эозинофилов. Продукция ИЛ-5 зависит от образования Тх2 из Тх0, а этот процесс контролируется ИЛ-4. Существует большое количество экспериментальных и клинических доказательств, что многие физиологические эффекты ИЛ-5 (например, его противопаразитарная защита) критически зависят от уровня ИЛ-4. В эксперименте было убедительно доказано, что мыши, дефицитные по ИЛ-4, более чувствительны к заражению паразитами и не способны изгонять взрослых червей Heligmosomoides при вторичном инфицировании.
Таким образом, ИЛ-4 является главным продуктом Тх2, стимулирует их дифференцировку, является антагонистом цитокинов, продуцируемых Тх1, влияет на развитие кроветворных клеток, на макрофаги, NK-клетки, базофилы. ИЛ-4 способствует развитию аллергических реакций, обладает противовоспалительным и противоопухолевым действием.
Возможные клинические применения ИЛ-4:
· Как возможный противовоспалительный агент при таких заболеваниях, как сепсис, ревматоидный артрит и др.;
· Как иммуномодулятор, подавляющий образование и функции Тх1, при лечении некоторых аутоиммунных заболеваний, например, диабет первого типа, острая РТПХ;
· Для местной противоопухолевой терапии.
ИНТЕРЛЕЙКИН-5 (кодируется у человека генами 5-й пары хромосом). Представляет собой сильно гликолизированный белок с молекулярной массой 45- 60 кД. Продуцируется Тх2. Основные клетки-мишени для ИЛ-5 – В-лимфоциты и эозинофилы.
Способность ИЛ-5 поддерживать пролиферацию В-клеток реализуется на более поздних этапах активации, чем действие ИЛ-4 и ИЛ-1. ИЛ-5, способствуя дифференцировке В-лимфоцитов в плазматические клетки, особенно в продуценты IgA, стимулирует выработку секреторного IgA, благодаря чему способствует формированию местной иммунной защиты слизистых оболочек.
В норме количество эозинофилов в крови невелико. В присутствии ИЛ-3 и ГМ-КСФ из миелоидных предшественников образуется лишь небольшое количество эозинофильных колоний. Однако при паразитарных инфекциях, например при гельминтозах, наблюдается резкое возрастание количества эозинофилов. То есть, ИЛ-5 является основным ростовым и дифференцировочным фактором эозинофилов, повышая их цитотоксичность по отношению к личинкам гельминтов. ИЛ-5 способствует вовлечению эозинофилов в воспалительные, антипаразитарные и противоопухолевые реакции. Кроме того, ИЛ-5 активирует зрелые эозинофилы: повышает их фагоцитарную функцию, усиливает антителозависимый цитолиз опухолевых клеток и продукцию супероксид-анион-радикалов кислорода.
Действие ИЛ-5 может быть усилено действием других интерлейкинов, например таких, как ИЛ-3 и ИЛ-4. Продукция ИЛ-5 напрямую зависит от продукции ИЛ-4 Т-хелперами-2. У мышей, дефицитных по ИЛ-4, резко снижена эозинофилия при паразитарных инфекциях, а у мышей, дефицитных по ИЛ-5, образование эозинофилов блокировано.
Таким образом, ИЛ-5 обладает достаточно “узким” спектром действия. Он является ростовым и дифференцировочным фактором В-лимфоцитов и эозинофилов, усиливает синтез IgA и тем самым способствует развитию местного иммунитета слизистых оболочек, стимулирует эзинофилы к участию в защите от паразитарных инвазий и опухолевого роста.
ИНТЕРЛЕЙКИН-6 (кодируется у человека генами 7-й пары хромосом). ИЛ-6 представляет собой гликопротеин с молекулярной массой 20-30 кД. Первоначально был открыт как лимфокин Т-клеточного происхождения, который индуцирует конечную стадию созревания В-лимфоцитов в плазматические клетки. В дальнейшем было доказано, что ИЛ-6 обладает многими видами биологической активности: секреция иммуноглобулинов В-клетками; синтез различных белков острой фазы в печени; обеспечение (облегчение) роста различных В-клеточных линий (миелома, плазмоцитома, гибридомы); созревание мегакариоцитов; дифференцировка нейронов; активация остеокластов. Клетки–продуценты ИЛ-6 очень многочисленны: моноциты/макрофаги, лимфоциты, фибробласты, гепатоциты, кератиноциты, эндотелиальные, мезангиальные, кроветворные клетки, клетки трофобласта и опухолей различного происхождения. Индукторами выработки ИЛ-6 могут быть бактериальные продукты, распознаваемые с помощью TLR и NOD-рецепторов врожденного иммунитета, полиэлектролиты, митогены, а также ИЛ-1, TNF-α, интерфероны и колониестимулирующие факторы.
Клетками-мишенями для ИЛ-6 являются: различные соединительно-тканные элементы, клетки крови, иммунной и нейроэндокринной систем, печени.
Биологические эффекты сходны с ИЛ-1 и TNF-α. Прежде всего это участие в воспалении, в индукции и реализации врожденного и адаптивного иммунного ответа, а также в кроветворении. ИЛ-6 способствует как обострению хронических, так и хронизации острых воспалительных процессов. Для него свойственно противовирусное действие. Выделяясь несколько позже, чем ИЛ-1 и TNF-α, ИЛ-6 подавляет их образование (эти цитокины, напротив, стимулируют его выработку) и поэтому относится к цитокинам, завершающим развитие воспалительной реакции.
В иммунной системе ИЛ-6 является фактором, стимулирующим пролиферацию В-лимфоцитов – их терминальной дифференцировки в плазматические клетки, продуцирующие антитела. Он равномерно стимулирует выработку иммуноглобулинов всех классов. Действуя на зрелые Т-клетки, ИЛ-6 подготавливает их к реакции на ИЛ-2, усиливает вызываемую ИЛ-2 пролиферацию Т-клеток и дифференцировку цитотоксических Т-лимфоцитов, повышает действие IFN-γ на NK-клетки.
На кроветворные клетки ИЛ-6 действует, в основном, как кофактор, способствуя проявлению эффектов ИЛ-3, ГМ-КСФ и М-КСФ. В гемопоэзе ИЛ-6 действует как костимулятор для ИЛ-3-зависимой пролиферации полипотентных стволовых клеток костного мозга и индуцирует созревание мегакариоцитов, что приводит к возрастанию количества тромбоцитов. Он может также стимулировать пролиферацию лейкемических миелоидных бластных клеток и кератиноцитов. ИЛ-6 выполняет определенную, но пока недостаточно ясную роль во взаимодействии иммунной и нейроэндокринной систем, влияя на развитие нейронов.
В связи с множественностью механизмов действия ИЛ-6 участвует в патогенезе многих заболеваний, включая СПИД, множественную миелому, ревматоидный артрит, постменопаузный остеопороз, способствует росту саркомы Капоши.
Таким образом, ИЛ-6 представляет собой полифункциональный цитокин, продуцируемый главным образом дендритными клетками, монцитами/макрофагами и другими клетками врожденного иммунитета. По спектру биологического действия он близок к ИЛ-1 и TNF-α, участвует в развитии воспаления, иммунных реакций, в регуляции кроветворения, служит ростовым фактором плазматических клеток, участвует в межсистемных взаимодействиях.
ИНТЕРЛЕЙКИН-7 (кодируется у человека генами 8-й пары хромосом).ИЛ-7 описан в 1988 г. как активатор предшественников лимфоцитов. Его молекулярная масса составляет 25 кД. Впервые был описан как фактор, стимулирующий развитие В-лимфоцитов. Затем было установлено, что ИЛ-7 является мощным стимулятором пролиферации предшественников Т- и В-лимфоцитов, действует как ростовой фактор для зрелых функциональных Т-клеток и способствует генерации литически активных антиген-специфических цитотоксических Т-лимфоцитов и ЛАК-клеток с помощью ИЛ-2-независимого механизма.
Продуцентами ИЛ-7 являются стромальные клетки костного мозга, тимуса (фибробласты, эндотелиальные клетки, эпителиальные клетки тимуса), селезенки, почки, а также макрофаги.
ИЛ-7 представляет собой основной лимфопоэтин. Он стимулирует развитие стволовых лимфоидных клеток в сторону В-лимфоцитов, стимулирует пролиферацию внутритимусных предшественников Т-лимфоцитов (мощный ростовой фактор для тимоцитов на ранних стадиях их развития), усиливает экспрессию ингибитора апоптоза bcl-2, способствуя выживаемости пре-Т-клеток, обусловливает антигеннезависимое размножение Т-лимфоцитов вне тимуса. Стимулируя пролиферацию тимоцитов, ИЛ-7 действует синергично с ИЛ-1 и в этом отношении ИЛ-7, играет центральную роль в развитии Т-клеток.
В качестве кофактора (подобно ИЛ-6) ИЛ-7 стимулирует продукцию ИЛ-2, влияя на пролиферацию Т-клеток и дифференцировку CD8+ ЦТЛ. Возможно, что ИЛ-7 может в какой-то мере замещать ИЛ-2 как митоген в генерации ЦТЛ и ЛАК-клеток.
В отличие от большинства других цитокинов, эффект которых дублируется, влияние ИЛ-7 на развитие лимфоцитов не имеет подобной страховки и удаление гена ИЛ-7 приводит к опустошению тимуса, развитию тотальной лимфопении и тяжелого иммунодефицита.
ИЛ-7 оказывает выраженное влияние на человеческие моноциты периферической крови: индуцирует в них секрецию больших количеств ИЛ-1α, ИЛ-1β, ИЛ-6 и TNF-α.
Таким образом, ИЛ-7, является лимфопоэтином, который играет важную роль в обеспечении Т-клеточного звена адаптивной иммунной системы.
ИНТЕРЛЕЙКИН-8 (кодируется у человека генами 4-й пары хромосом). Относится к семейству хемокинов. Его молекулярная масса составляет 75 кД.
Продуцентами ИЛ-8 являются моноциты/макрофаги, Т-лимфоциты, а также фибробласты, гепатоциты, нейтрофилы, клетки глиобластомы и астроцитомы, NK-клетки и ряд других клеток. Стимуляторами продукции ИЛ-8 являются: фитогемагглютинин, конканавалин А, ЛПС, ИЛ-1α и ИЛ-1β, TNF-α, ИЛ-3, IFN-γ, форболовые эфиры, вирусы и бактерии.
Основная мишень для ИЛ-8 – это нейтрофилы, на хемокиновые рецепторы которых он действует как хемоаттрактант и активатор (хемотаксис, изменение формы; дегрануляция, дыхательный взрыв, продукция супероксида; повышение концентрации Са+2 в цитозоле; усиление адгезии к клеткам эндотелия, фибриногену и белкам внеклеточного матрикса; ингибирование роста Саndida albicans; усиление выделения лизосомальных ферментов). Кроме того, ИЛ-8 обеспечивает хемотаксис Т-лимфоцитов и базофилов в очаг реакции, совместно с ИЛ-4 стимулирует дегрануляцию тучных клеток и базофилов, усиливая выделение гистамина и лейкотриенов. Повышает адгезию моноцитов к клеткам эндотелия, усиливает секреторную активность макрофагов, вызывает хемотаксис эозинофилов, повышает адгезивность клеток меланомы, стимулирует рост кровеносных сосудов.
Таким образом, ИЛ-8 является одним из представителей семейства цитокинов, которые индуцируются при воспалении и действуют, в основном, как хемокины для целого ряда клеток. В случае с ИЛ-8 такими клетками прежде всего являются нейтрофилы. Поэтому выделение ИЛ-8 из макрофагов воспалительных очагов или из тканевых макрофагов специфически привлекает нейтрофилы в зону повреждения или инфекции.
ИНТЕРЛЕЙКИН-9 (кодируется у человека генами 5-й пары хромосом, локализованными недалеко от гена для ИЛ-3). Является фактором роста активированных клонов Т-хелперов, а также тучных клеток. Имеет молекулярную массу 30-40 кД. ИЛ-9 синтезируется, в основном, Тх2 (при стимуляции лектинами и антителами против CD3) позже других цитокинов.
|
|
|
|
|
Дата добавления: 2014-11-25; Просмотров: 2109; Нарушение авторских прав?; Мы поможем в написании вашей работы!