КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Анатомические и физиологические основы гистопатологических и электрофизиологических исследований 1 страница
|
|
|
|
Моторная единица служит окончательным общим трактом для проведения моторной активности в нервной системе, а мышца является окончательным эффекторным органом моторной единицы. Все движения человеческого тела, его положение в пространстве и рефлекторная активность — это результат интегрированных импульсов большого числа моторных единиц, опосредованных как спинально, так и супраспинально. Сила мышечного сокращения зависит от числа моторных единиц, участвующих в данном сокращении; частоты, с которой происходит разрядка импульсов в моторной единице; скорости сокращения мышечных волокон в моторной единице и от природы (характера) моторной единицы (является ли она устойчивой к утомлению или, напротив, склонной к нему). Число моторных единиц в различных мышцах различно и колеблется от 100 в мышцах кисти руки до нескольких тысяч в мышцах ног. Число мышечных волокон, приходящихся на моторную единицу, также колеблется от нескольких (около 10) в наружных мышцах глаза до почти 2000 в мышцах ног, например в икроножной мышце. Число же мышечных волокон в одной мышце колеблется от 1000 в наружных мышцах глаза до 1 млн и более в больших мышцах ног. Чтобы правильно интерпретировать результаты клинических и лабораторных исследований как нормальной, так и поврежденной мышцы, необходимо понимать организацию моторных единиц и характер их возбуждения. Мышечные волокна моторной единицы распределены в мышце по случайному принципу и волокна, иннервируемые одной и той же клеткой переднего рога спинного мозга, как правило, не соприкасаются. Моторные единицы различаются между собой как по размерам, так по биохимическим и физиологическим свойствам их мышечных волокон. На основании этих свойств, выявляемых при окрашивании гистологических препаратов мышц на АТФазу, мышечные волокна подразделяют на два типа. Тип I — это волокна, которые прокрашиваются лишь незначительно и тип II — это волокна, которые в препарате выглядят темными. Названный метод гистологической идентификации имеет большую диагностическую значимость, поскольку некоторые мышечные заболевания характеризуются преобладанием патологических изменений в мышечных волокнах лишь одного типа. Так, число волокон II типа уменьшается при врожденных миопатиях, в то время как при миотонической дистрофии атрофии подвергаются именно волокна I типа.
По-видимому, все мышечные волокна, относящиеся к данной моторной единице, принадлежат к одному гистохимическому типу. Эксперименты на животных по перекрестной иннервации, у которых все волокна данной мышцы относятся к одному гистохимическому типу (тогда как в мышцах человека нервные волокна разных типов распределены в данной мышце по случайному принципу), показали, что гистохимические, биохимические и физиологические свойства мышечных волокон меняются в ответ на изменение иннервации. Основой для контроля свойств мышечного волокна со стороны нервной системы, вероятно, являются особенности импульсного разряда, поскольку при экспериментальной хронической электрической стимуляции нервов биохимические и физиологические свойства мышцы меняются. Подобное же изменение типа мышечного волокна у человека может быть вызвано длительным упражнением данной мышцы; при этом увеличивается число волокон I типа (аэробного).
Физиологические свойства мышечных волокон определяют способность мышцы выполнять различные физические нагрузки. Моторные единицы с мышечными волокнами I типа или мышечные волокна с реакцией медленного подергивания осуществляют длительную и продолжительную активность, так как их энергетическое снабжение за счет продукции АТФ основано на метаболизме субстрата, происходящего благодаря окислительным процессам в митохондриях. Мышечные волокна этих моторных единиц подчинены небольшим, низкопороговым моторным нейронам с медленным проведением импульса, активизируемого напряжением небольшой интенсивности. Усилие же большой интенсивности, например поднятие тяжелого груза, требует включения больших высокопороговых моторных нейронов с быстрым проведением импульса, которые иннервируют II тип мышечных волокон.
Физиологические свойства моторных единиц и их ответная реакция на произвольные мышечные сокращения таковы, что существует стереотипный характер их мобилизация для каждой мышцы со строго определенной последовательностью активации мышечных единиц. Некоторые моторные единицы активизируются только большой активностью. Это частично обусловливает способность мышцы восстанавливать свой размер и силу при повторных значительных напряжениях. При этом число миофибрилл в мышечных волокнах увеличивается, а возможно, в какой-то степени увеличивается и число самих мышечных волокон. Недостаточная активация некоторых моторных единиц в обычных условиях и их включение при очень сильных тренировках может объяснить мышечную слабость, столь обычную у многих лиц, ведущих сидячий образ жизни, а также положительное влияние умеренных физических нагрузок на таких людей.
По мере расслабления мышцы прекращение поступления импульсов в отдельные моторные единицы происходит по отдельным группам волокон, так что лицо, производящее неадекватное мышечное напряжение, вследствие функциональной слабости (например, при истощении), в случае отсутствия мотивации или при болевом ощущении, часто выполняет его по типу «зубчатого колеса» или по типу «дай дорогу», что позволяет различить истинную и симулированную мышечную слабость.
Электромиография
Измерение электрической активности, возникающей в мышечных волокнах, осуществляют введением иглы электрода чрескожно в мышцу. Электрическая активность с этого электрода затем визуализируется на катодно-лучевом осциллоскопе и может быть трансформирована в звуковой сигнал.
Скелетные мышцы нередко имеют очень большие размеры. Поэтому электродные исследования таких мышц дают лишь усредненную картину их электрической активности. Поскольку при нервно-мышечных заболеваниях поражается только скелетная мускулатура, нормальная электрическая активность в одном из участков мышцы не исключает возможности патологического процесса в соседнем ее участке. Таким образом, для тщательного физиологического анализа необходимы тонкие концентрические игольчатые электроды, которые должны быть введены в строго определенные мышцы, для того чтобы регистрировать электрическую активность небольшого числа моторных единиц и мышечных волокон.
Потенциал действия. По мере того как электрический импульс распространяется от центра к любому концу мышечного волокна, электрический ток проводится на поверхности через нормально поляризованную область мышечной мембраны (сарколеммы) по направлению к деполяризованной ее зоне (рис. 354-1). Регистрирующий электрод вначале приобретает слегка положительный заряд по отношению к индифферентному электроду. Когда зона деполяризации движется под регистрирующим электродом, то на электромиограмме (ЭМГ) появляется негативный зубец. По мере того как активная зона мышцы удаляется от электрода, мембрана под ним постепенно реполяризуется. В результате на осциллоскопе записывается трехфазный потенциал действия (см. рис. 354-1).
Регистрация активности моторной единицы (двигательной единицы). Трехфазные потенциалы действия образуются в результате активности одиночных мышечных волокон. В нормальной мышце возбуждение начинается с возбуждения моторных нервов, иннервирующих многие мышечные волокна, так что все мышечные волокна одной данной моторной единицы активизируются одним импульсом, передающимся с моторного нейрона. Число мышечных волокон в одной моторной единице различно, а мышечные волокна отличаются друг от друга диаметром, длиной и формой, а также ориентацией в пространстве по отношению к электроду электромиографа. Мышечные волокна одной моторной единицы не собраны в пучки, а довольно широко разбросаны в том или ином участке мышцы, будучи перемешанными с мышечными волокнами расположенных рядом моторных единиц. Следовательно, активизация мышцы сопровождается образованием целого комплекса потенциалов моторных единиц, являющихся суммой индивидуальных потенциалов действия.
Нормальная электромиограмма. В покое нормальная мышца не несет электрического заряда, она электрически «молчалива». По мере того как замирает ее активность. индуцированная введением игольчатого электрода, т. е. активность, спровоцированная травмой от укола, электроды уже не регистрируют никаких потенциалов действия. Когда же данная мышца произвольно сокращается, то появляются потенциалы действия, причем по мере усиления силы сокращения мышцы включаются единицы
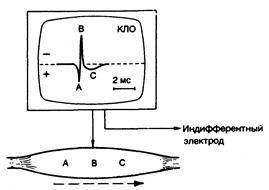
Рис. 354-1. Трехфазный потенциал действия мышцы. Затененные участки представляют зону потенциала действия, которая заряжена отрицательно по отношению ко всем другим точкам на поверхности мышечного волокна. Это обозначено тремя точками (А, В, С) по движению потенциала вдоль мышечного волокна. В каждой точке часть трехфазного мышечного потенциала действия отображается на катодно-лучевом осциллоскопе (КЛО), где регистрируется разница потенциалов между активным (вертикальная стрелка) и индифферентным электродами. На этом и последующих рисунках отрицательная полярность изображена вверху. Время откалибровано на экране КЛО. (Подробности см. в тексте.)
второго и третьего порядка. При усилении мышечного сокращения появляются потенциалы действия еще большего числа и более крупных моторных единиц, пока уже при полном сокращении мышцы не регистрируются беспорядочно расположенные потенциалы действия, частота которых колеблется от 20 до 50 Гц. При этом потенциалы действия моторных единиц больше уже не различаются и может быть получена электромиограмма периода «полной мобилизации», или фазы интерференции. При заболеваниях центральной или периферической нервной системы этот период мобилизации, возникающий после максимального волевого усилия, направленного на сокращение мышцы, резко укорочен, так как при этом активизируется меньшее число моторных единиц. У больных же с поражением мышц период мобилизации при совершении максимального усилия остается полным; однако «peak-to-peak» амплитуда фазы «мобилизации» уменьшена.Таким образом, уменьшение амплитуды фазы полной «мобилизации» при слабости мышцы, выявляемое при ЭМГ, служит самым характерным электромиографическим признаком мышечного заболевания.
Патологическая электромиограмма. Нарушения, регистрируемые на ЭМГ, характеризуются: 1) возникновением спонтанной активности во время релаксации мышцы (фибрилляции, положительные волны, фасцикуляции); 2) различными отклонениями в амплитуде, продолжительности и форме отдельных потенциалов моторных единиц;
3) уменьшением числа моторных единиц, которое, однако, может пополняться; 4) обнаружением миотонии, парных потенциалов (тетания), странных повторяемых потенциалов или электрического «затишья» во время укорочения (сокращения) мышцы.
Спонтанная активность во время полного расслабления мышцы. Персистирующую инсерционную (от введения в мышцу игольчатого электрода) активность можно наблюдать при нарушениях мышечного тонуса, при полимиозите и в денервированных мышцах. Спонтанную активность всей моторной единицы или ее части называют фасцикуляцией, а спонтанную активность отдельного мышечного волокна — фибрилляцией. Фибрилляции возникают при деструкции моторного нейрона или его аксона, а также при мышечных заболеваниях, конца часть мышечного волокна отделяется от его иннервируемой части очагом сегментарного некроза. Когда моторный нейрон подвергается деструкции в результате патологического процесса или когда его аксон разрывается, дистальная часть аксона в течение нескольких дней подвергается дегенерации. Мышечные волокна, которые в прошлом иннервировались веточками погибшего аксона, как бы отъединяются от нервной системы. Хемочувствительная часть сарколеммы в области моторной концевой пластинки после денервации начинает распространяться на всю поверхность мышечного волокна. Это наряду со снижением мембранного потенциала покоя денервированного мышечного волокна вызывает появление спонтанной активности в денервированных мышечных волокнах в период 7—25 дней после «смерти» аксона (время определяется расстоянием от денервированных мышечных волокон до участка поражения). Эта спонтанная активность подобна той, которую обнаруживают в синусовом узле сердца, т. е. каждое мышечное волокно сокращается со своей собственной скоростью безотносительно активности мышечных волокон, расположенных рядом. При этом на ЭМГ денервированная мышца может давать совершенно случайный конгломерат потенциалов, состоящий из коротких, трехфазных фибрилляционных потенциалов и двухфазных положительных острых волн. Фибрилляции и активность положительных острых волн обнаруживают до тех пор, пока мышечные волокна не подвергнутся реиннервации за счет разрастания новых аксонов из проксимального конца поврежденного нерва или из волокон расположенного рядом здорового нерва, или пока, наконец, спустя месяцы и годы мышечные волокна не подвергнутся замене соединительной тканью. Фасцикуляции — это непроизвольные единичные сокращения всей моторной единицы или ее части. Поскольку огромное число мышечных волокон сокращается одновременно, то возникает видимая на глаз дефигурация кожи, покрывающей мышцу (образование ямочки или подергивание), однако при этом нет достаточной силы, чтобы вызвать движение в суставе, относящемся к данной мышце. Форма возникающих при этом на ЭМГ потенциалов, как и таковых обычной моторной единицы, относительно постоянна для каждой данной фасцикулирующей единицы. При этом наблюдают полифазную кривую, состоящую из трех—пяти фаз продолжительностью 5—15 мс, амплитуда которых несколько сот микровольт. При доброкачественной фасцикуляции, отмечаемой у многих нормальных лиц, одна и та же единица имеет тенденцию сокращаться с регулярной скоростью, что указывает на ритмическую стимуляцию мышечных волокон, иннервируемых данным аксоном. Изменения на ЭМГ при доброкачественных фасцикуляциях практически соответствуют потенциалам нормальной моторной единицы. Если же частота разрядов превышает таковую при фасцикуляциях, это свидетельствует о каком-либо нарушении фасцикуляции, отмечаемом при медленно прогрессирующих заболеваниях клеток переднего рога, как, например, при амиотрофическом боковом склерозе и при прогрессирующей спинальной мышечной атрофии, обычно многочисленны, относительно пролонгированы, с высокой амплитудой. Такие же фасцикуляции появляются при компрессионных поражениях нервного корешка, при некоторых моторных невропатиях и на ранних этапах острой воспалительной полиневропатии. В случае поражения нервных корешков, как это бывает при грыже Шморля (с разрывом волокон диска), повреждается большое число аксонов, что сопровождается резко выраженными фасцикуляциями. В подобных ситуациях поврежденный нейрон как бы «раздражен» патологическим процессом, разряды импульсов в нем происходят многократно, и при этом возникает активность в мышечных волокнах, иннервируемых данным нейроном.
Фасцикуляции могут быть спровоцированы травматическими повреждениями периферического нерва, после смерти аксона их сменяют фибрилляции. Фасцикуляции в икроножных мышцах и мышцах кисти часто возникают и у здоровых лиц.
Патологические изменения в потенциалах моторной единицы. Патологические изменения потенциалов моторной единицы отражаются на их амплитуде, числе, продолжительности и форме.
Увеличение амплитуды. Через короткое время после денервации моторные единицы, находящиеся в функциональной связи со спинным мозгом, остаются нормальными, но число потенциалов моторных единиц, обнаруживающихся во время мышечного сокращения, уменьшается. Поздние потенциалы оставшихся моторных единиц также претерпевают изменения: увеличивается их амплитуда, превышая норму в 2—3 раза, возрастает продолжительность, они становятся полифазными (более четырех фаз). В моторных единицах, содержащих большое число мышечных волокон, возникают очень большие потенциалы (более 5 мВ) (рис. 354-2, в), причем эти мышечные волокна в данной мышце располагаются на большей площади. Подобные увеличенные моторные единицы появляются в тех случаях, когда отростки новых веточек из неповрежденных аксонов вновь иннервируют (реиннервируют) ранее денервированные мышечные волокна, тем самым присоединяя их к своим собственным моторным единицам. Эти реиннервированные моторные единицы подчас генерируют уже полифазные и пролонгированные потенциалы действия, что является патогномоничным для феномена реиннервации. Эти потенциалы должны быть дифференцированы от полифазных потенциалов нормальной продолжительности, которые могут возникать в нормальных мышцах,
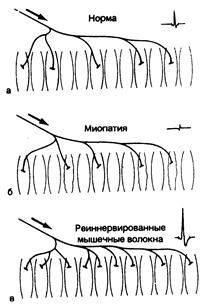
Рис. 354-2. Потенциалы моторной единицы.
Заштрихованные мышечные волокна — функциональные участки одной моторной единицы. Аксон (направление его показано стрелкой) разветвляется на терминальные волокна, иннервирующие соответствующие мышечные волокна; потенциал действия моторной единицы, образуемый каждой из них, показан справа вверху; его продолжительность определяется промежутком между двумя небольшими вертикальными линиями. Незаштрихованные мышечные волокна относятся к другим моторным единицам. а — норма; изображено пять мышечных волокон в активной (моторной) единице; б — в этой миопатической моторной единице только два мышечных волокна остались активными; остальные три (с неровными контурами) поражены мышечным заболеванием; в — четыре мышечных волокна, относящиеся к другим моторным единицам и которые были денервированы ранее, в настоящее время вновь реиннервированы ответвлением терминальной веточки аксона, принадлежащего здоровой моторной единице. Как моторная единица, так и ее потенциал действия теперь представляются большими, чем в норме. Обратите внимание, что только при этих ненормальных условиях мышечные волокна в одной и той же (моторной) единице расположены рядом друг с другом.
особенно в области концевых пластинок, а также от коротких полифазных потенциалов, характерных для патологии мышц.
Уменьшение амплитуды и продолжительности потенциала. При таких болезнях, как полимиозит, мышечные дистрофии и другие миопатии, при которых разрушаются рассеянные по моторной единице мышечные волокна (см. рис. 354-2, б), а также уменьшается и популяция мышечных волокон, приходящихся на одну моторную единицу. Когда подобная единица активизируется, амплитуда ее потенциалов снижена, а продолжительность уменьшена по сравнению с нормой; такой потенциал может казаться полифазным, поскольку видны потенциалы отдельных мышечных волокон. Когда большинство мышечных волокон данной моторной единицы поражено, потенциалы действия трудно отличить от фибрилляций, а когда деструктивный процесс охватывает все мышечные волокна, электрическая активность мышцы прекращается. Небольшие непродолжительные потенциалы действия вызывают характерное потрескивание высокого тона в звуковом регистраторе электромиографа. При миастении и при других заболеваниях, сопровождающихся прогрессирующим нарушением передачи нервных импульсов во все новых и новых нервно-мышечных соединениях, электромиографический потенциал данной моторной единицы сначала может быть нормальным, но по мере развития мышечного утомления становится «миопатическим». Потенциалы действия с ослабленных мышц при миастении представляются пропорционально «миопатическими». Как показано на рис. 354-2, б. потенциалы моторной единицы выглядят равно «миопатическими», если патологическим процессом или прямо поражаются отдельные мышечные волокна в пределах моторной единицы, как это бывает при мышечной дистрофии, или же нарушается нервно-мышечная передача нервного импульса в каком-нибудь одном нервно-мышечном соединении, как при миастении.
Уменьшение числа моторных единиц. Заболевания, при которых уменьшается число нижних моторных нейронов или моторных аксонов в периферическом нерве, приводят к уменьшению числа моторных единиц, которое может быть восполнено в пораженных мышцах. Число моторных единиц, доступных активации в денервированных мышцах, варьирует пропорционально силе максимального произвольного сокращения мышцы, а потенциал действия на ЭМГ уже не выглядит интерферентным, но скорее похож на потенциал отдельной моторной единицы или при максимальном волевом усилии носит смешанный характер.
Уменьшение числа мышечных волокон. При таких мышечных заболеваниях, как мышечная дистрофия или другие миопатии, когда поражаются отдельные мышечные волокна, как правило, очень незначительно (или этого не происходит совсем) уменьшается число моторных единиц, которые могут быть использованы для восполнения утраченных, хотя каждая единица при этом содержит меньшее число мышечных волокон, чем в норме. При максимальном произвольном усилии на ЭМГ регистрируется кривая, характерная для полной мобилизации, несмотря на выраженную слабость мышцы. Но в связи с тем что активизируется меньшее число мышечных волокон, амплитуда кривой уменьшена по сравнению с нормой. Таким образом, кривая в виде «полной мобилизации», но уменьшенной по сравнению с нормой амплитуды при наличии клинически значительной мышечной слабости является характерной чертой так называемой миопатической электромиограммы.
Варианты форм потенциалов действия. При некоторых поражениях нервно-мышечного соединения, как, например, при миастении или при синдроме Ламберта—Итона, образуются патологические формы и амплитуды потенциалов отдельных моторных единиц. Эти колебания потенциалов моторной единицы являются результатом перемежающейся по интенсивности блокады проведения в отдельных нервно-мышечных соединениях.
Другие нарушения, выявляемые на ЭМГ. При миотонии сарколеммные мембраны очень легко раздражимы, так что повторная мышечная деполяризация и мышечное сокращение возникают, несмотря на усилия больного расслабить мышцу (см. гл. 17). Такой характер ЭМГ приобретает при врожденной миотонии, миотонической дистрофии и гиперкалиемическом периодическом параличе. Миотония сопровождается образованием на ЭМГ высокочастотных повторных разрядов, амплитуда и частота которых то усиливается, то ослабевает, а в звуковом регистраторе появляется звук «движущегося мотоцикла» или «ныряющего бомбардировщика». Феномен миотонии на ЭМГ возникает при перкуссии мышцы, при движении игольчатого электрода или, наконец, вслед за произвольным сокращением мышцы. Потенциалы моторной единицы выглядят как будто и нормально, однако за ними не следует периода электрического покоя, который в норме наступает после расслабления мышцы. Вместо этого наблюдают «взрыв» быстрой активности, который может продолжаться даже в течение нескольких минут и только потом затихает. Некоторые потенциалы этого удлиненного разряда имеют продолжительность, амплитуду и форму активности одиночного мышечного волокна, в то время как другие похожи на потенциалы моторных единиц. При гипотиреозе и другой патологии, поражающей периферические нервы или мышцы, на ЭМГ видны уродливые высокочастотные повторные разряды без волн усиления или ослабления. Высокочастотные парные потенциалы действия, складывающиеся из двойных, тройных и более кратных одиночных (моторных) единиц, возникают при тетании и «спазме половины лица» и указывают на нестабильность реполяризационного процесса нервного волокна. При контрактурах, как, например, при болезни Мак-Ардла или при злокачественной гипертермии, электрических потенциалов нет.
ЭМГ одиночного мышечного волокна и макро-ЭМГ. Помимо обычной электромиограммы (ЭМГ) с концентрическими игольчатыми электродами, специальная техника позволяет регистрировать ЭМГ отдельных мышечных волокон или сразу всей моторной единицы (макро-ЭМГ). Посредством техники ЭМГ, регистрирующей потенциалы отдельного мышечного волокна, можно регистрировать также «джиттер» (вздрагивание) и измерять с точностью до микросекунд процессы, происходящие в отдельных нервно-мышечных соединениях. При этом характерные количественные отклонения от нормы обнаруживают при миастении и при других нарушениях нервно-мышечных соединений.
Исследования с одноволоконными ЭМГ используют для расчета волоконной плотности — число отдельных одноволоконных потенциалов действия, принадлежащих к одной моторной единице в пределах регистрируемого участка одноволоконного ЭМГ-электрода (приблизительно 200 мкм). Величина волоконной плотности возрастает после денервационно-реиннервационных процессов.
Посредством техники макро-ЭМГ регистрируют потенциалы всех мышечных волокон, принадлежащих к моторной единице; она позволяет оценить истинные ее размеры. Амплитуда и площадь макро-ЭМГ, регистрирующей потенциалы моторной единицы, увеличиваются при реиннервации и уменьшаются при первичных мышечных заболеваниях, сопровождающихся уменьшением числа мышечных волокон, приходящихся на одну моторную единицу.
Исследования нервной проводимости
Стимулирование достаточно больших моторных и чувствительных нервов позволяет регистрировать их потенциалы действия и получать объективные количественные данные относительно латентного периода и скорости проведения импульса по нерву. Методика основана на стимулировании нерва поверхностными электродами, помещаемыми на кожу над исследуемым нервом. В результате электроды регистрируют составной потенциал действия в нерве, расположенном проксимально (при исследовании больших сенсорных волокон) или над мышцей, расположенной дистально (в случае исследования моторных волокон в смешанном двигательно-чувствительном нерве) (рис. 354-3). Время проведения импульса от наиболее дистально расположенного стимулируемого электрода, измеренное в миллисекундах, с момента стимуляционного воздействия до начала ответной реакции, получило название дистального, или периферического латентного, времени. Если второй стимул наносится на смешанный нерв более проксимально (или если регистрирующие электроды расположены более проксимально в случае исследования чувствительных волокон), то можно измерить новое и более продолжительное время проведения. Если расстояние (в миллиметрах) между двумя участками стимуляции двигательных волокон или регистрации с чувствительных волокон разделить на разницу показателей времени проведения (в миллисекундах), можно получить максимальную скорость проведения (в метрах в секунду). Она показывает скорость распространения потенциалов действия в наиболее крупных и наиболее быстро проводящих импульс нервных волокнах. Показатели этих скоростей у здоровых лиц колеблются довольно значительно — от 40—45 м/с (в зависимости от исследуемого нерва) до 75—80 м/с. У новорожденный эти значения несколько ниже (они составляют примерно 1/2 от показателей взрослых), но достигают данного уровня к 3—4 годам жизни. Получены показатели нормы периферических латентных периодов для наиболее дистальных участков различных смешанных нервов, иннервирующих соответствующие мышцы. Например, когда стимулируют срединный нерв у запястья, латентный период для проведения импульса через канал запястья к короткой отводящей мышце большого пальца у здоровых лиц составляет менее 4,5 м/с. Составлены специальные таблицы с подобными нормативами для скорости проведения и дистальных латентных периодов, показатели которых колеблются в зависимости от расстояния. Во время исследования нервной проводимости очень важно поддерживать нормальную температуру тела человека, так как при субнормальной температуре скорость проведения по нерву замедляется. Скорость проведения по нерву зависит от диаметра нервного волокна и степени его демиелинизации. Немиелинизированные нервные волокна малого диаметра характеризуются меньшей скоростью проведения, чем миелинизированные волокна большого диаметра. У волокон с сегментарной демиелинизацией скорость проведения, как правило, уменьшена. При стимулировании моторных волокон периферического нерва при условии, что каждое нервное волокно находится в функциональном единении с иннервируемыми им многочисленными мышечными волокнами, с кожного электрода над исследуемой мышцей можно зарегистрировать составной мышечный потенциал действия, являющийся результатом импульсных разрядов многих мышечных волокон. Сенсорные потенциалы действия, регистрируемые непосредственно с самих нервных волокон, не обладают качеством «амплификации», создаваемым мышечными волокнами; дело в том, что для этого требуется большая электронная амплификация. В случае патологии нервов сенсорные потенциалы могут быть небольшими или их может не быть вообще, и, таким образом, зарегистрировать сенсорную проводимость становится невозможно. Напротив, достаточно надежно измерить скорость моторной проводимости возможно, даже если сохранным осталось лишь одно мышечное волокно. Измерения скорости нервной проводимости отражают
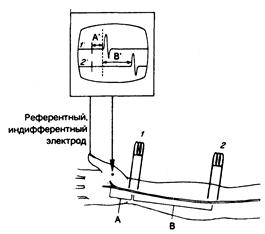
Рис. 354-3. Измерение скорости проводимости по нерву. Срединный нерв раздражают через кожу на уровне запястья (1) или в передней локтевой ямке (2), в результате чего возникает составной мышечный потенциал действия, регистрируемый как разница потенциалов между поверхностным электродом над выпуклостью тенара (вертикальная стрела) и индифферентным (референтным) электродом (РЕФ), расположенным более дистально. Изгиб кривой 1' на катодно-лучевом осциллоскопе (КЛО) отражает момент воздействия раздражения в точке 1, за которым следует мышечный потенциал. Дистальная «латентность» — это время А' на кривой КЛО (например, 3 м/с), которая соответствует проведению импульса на отрезке А по кисти.
То же самое относится и к кривой 2' (стимул воздействует в точке 2, а время от начала его воздействия до ответной реакции равно А' +В'). Максимальную скорость моторного проведения от точки 2 до точки 1 можно получить, разделив расстояние В на время В'.
|
|
|
|
|
Дата добавления: 2014-11-20; Просмотров: 594; Нарушение авторских прав?; Мы поможем в написании вашей работы!