КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Два принципа построения движения
|
|
|
|
Управление двигательными актами строится на двух основных принципах — принципе сенсорных коррекций текущего движения по цепи обратной связи и принципе прямого программного управления. Последний особенно важен для тех случаев, когда имеются быстрые изменения в системе, ограничивающие возможность сенсорных коррекций.
Накоплено множество экспериментальных фактов, подтверждающих реальное существование двух механизмов управления движением; посредством центральных моторных программ и с помощью обратной афферентации, которая используется для непрерывного контроля и коррекции выполняемого движения. Для многих

тттт-п-
Вспышки
Вспышки, прекращенные
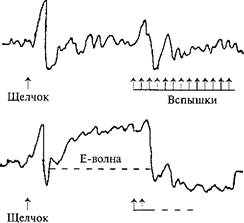
Рис. 52. Волна ожидания при отведении ЭЭГ от вертекса.
1 и 2 — реакции на изолированное действие щелчка и вспышек света; 3 —на комбинированное действие обоих стимулов, следующих один за другим; 4 — Е-волна (ожидания), когда испытуемый должен был нажимать на кнопку при появлении вспышек света и тем самым их прерывать. Интервал между щелчком (предупреждающим стимулом) и вспышками (императивным стимулом) фиксирован. Видна медленная негативность (отклонение вверх), предшествующая появлению императивного стимула (по Г. Уолтеру, 1966).
видов движения управление может осуществляться одновременно двумя механизмами при разном их соотношении для движений, различающихся сложностью и уровнем организации
Существуют два типа командных нейронов. Одни из них лишь запускают ту или другую двигательную программу, но не участвуют в ее дальнейшем осуществлении. Это нейроны-тригеры. Приме-
ром такого нейрона является маутнеровская клетка рыбы. Командные нейроны другого типа получили название воротных нейронов. Они поддерживают или видоизменяют двигательные программы, лишь будучи постоянно возбужденными. Такие нейроны обычно управляют позными или ритмическими движениями. В качестве примера можно привести нейрон генератора локомоторного ритма у таракана.
На принципиальную роль афферентации в регуляции движения и поведения в целом указывали НА. Бернштейн (1966) и П.К. Анохин (1968). Сенсорные раздражения не только могут запускать движение, но и выполнять корректирующую функцию. Обратная аффе-рентация, сигнализирующая о результатах действия, сопоставляется с программой движений и служит уточнению координат цели и траектории движения.
Значение афферентных систем для контроля за мотонейронами спинного мозга впервые было показано английским физиологом Ч. Шеррингтоном. Он указал на существование сенсорных обратных связей, которые регулируют активность мотонейрона. Им введен термин «.проприоцепция» для обозначения сенсорных входов, которые возбуждаются во время движения. Ч. Шеррингтон считал, что главная функция проприоцепторов — давать информацию о собственных движениях организма. Выделено два типа проприоцепторов; рецепторов растяжения, активирующихся при растяжении мышц, и рецепторов напряжения, чувствительных к силе сокращения мышц.
На основе опытов с условными рефлексами И.П. Павлов установил, что моторная кора получает сенсорные проекции от рецепторов мышечной и суставной чувствительности. В 1909 г. он ввел понятие «двигательный анализатор», выполняющий функцию восприятия сигналов от тела. Позже это понятие было расширено: в него были включены сенсорные и ассоциативные зоны коры, которые проецируются на моторную кору. В результате моторная кора стала рассматриваться как центральный аппарат построения движения.
Принципиальная невозможность реализации особенно произвольного движения с помощью одних только эфферентных импульсов была показана Н.А. Бернштейном. Он писал, что «...двигательный эффект центрального импульса не может быть предрешен в центре, а решается целиком на периферии... решающую роль для осуществления управления движением должна играть афферентация» (1966. С. 214). Согласно Н.А. Бернштейну, изменения в мышце, возникающие при движении, возбуждают чувствительные окончания проприоцепторов, а эти проприоцептивные сиг-
налы, достигая моторных центров, вносят изменения в эффектор-ный поток, т.е. в физиологическое состояние мышцы. Таким образом, перед нами «...не рефлекторная дуга, а другая форма взаимоотношений между афферентным и эффекторным процессами, характерная для всех координационных процессов, — рефлекторное кольцо» (1966. С. 91). Рефлекторное кольцо представляет собой фундаментальную форму течения двигательного нервного процесса.
Значение афферентации для осуществления движения становится очевидным при некоторых неврологических нарушениях, получивших название «апраксий». Под ними понимают-нарушения произвольных движений и действий, совершаемых с предметами. При кинестетической апраксий нарушается проприоцептив-ная кинестетическая афферентация. Патология возникает в задних отделах коркового ядра двигательного анализатора (в полях 1, 2 и частично 4). У такого больного движения становятся недифференцированными, плохо управляемыми, страдает письмо. При этом остается сохранной внешняя пространственная организация движения. Дефекты движения, возникающие из-за нарушений про-приоцептивной афферентации, частично могут быть компенсированы за счет усиления зрительного контроля. При пространственной апраксий поражение захватывает теменно-затылочные отделы коры на границе полей 19 и 39, нарушается зрительно-пространственная афферентация движений. Больные не могут выполнять пространственно-ориентированных движений (застелить постель, одеться). Усиление зрительного контроля не улучшает выполнение произвольных движений и действий, совершаемых с предметами.
До недавнего времени мало что было известно о процессах формирования и построения новых моторных программ. Однако благодаря изучению нейронной активности коры больших полушарий и других структур мозга у высших животных, а также клиническим данным, полученным при наблюдении за человеком, сложилось мнение, что ведущая роль в построении новых моторных программ принадлежит лобным отделам коры больших полушарий (префронтальной коре). Поражение в префронтальной коре может привести вообще к распаду произвольной регуляции движений. Этот дефект особенно выражен, когда требуется менять программы движений. Передние отделы коры для построения новых моторных программ используют весь видовой и накопленный в течение жизни индивидуальный опыт. При этом, по-видимому, происходит выбор отдельных фрагментов из хранящихся в памяти программ (как для врожденных форм поведения, так и приобретенных двигательных автоматизмов) для их последующей интеграции в новую моторную программу.
16-3341
Роль передних отделов коры больших полушарий в программировании движения руки обезьяны изучалась А. С. Батуевым- Им было обнаружено три группы нейронов, последовательно вовлекаемых в процесс осуществления условнорефлекторного двигательного навыка. Все три популяции нейронов были зарегистрированы в теменной и особенно в лобной коре.
Первая группа клеток реагировала только на условный сигнал и при этом по-разному на зажигание лампы слева и справа, т.е. эти нейроны реагировали на пространственные свойства условного раздражителя. Они были отнесены автором к сенсорным нейронам с пространственной селективностью. Вторая группа нейронов активировалась только в период отсрочки, обнаруживая селективность к условному сигналу слева и справа. Третья группа нейронов, обладавших также свойством пространственной селективности, отвечала лишь после открытия экрана, т.е. в пусковой период программы.
Исследователи предполагают, что сенсорные нейроны (первая группа) передают информацию второй группе нейронов, принадлежащей кратковременной памяти, которые возбуждаются в течение всего периода отсрочки, сохраняя тем самым полученную информацию. Третья группа нейронов — нейроны моторных программ, вероятно, получают информацию от нейронов памяти и запускают хорошо отработанную двигательную реакцию нажатия на правый или левый рычаг. Таким образом, нейроны передних отделов коры мозга принимают непосредственное участие в построении сложного двигательного акта, отвечающего требованиям пространственно-временных характеристик среды. А.С. Батуев говорит о последовательном эстафетном вовлечении различных популяций клеток в процесс программирования нового двигательного акта (Батуев А.С., 1991).
Программирующая функция префронтальной коры исследовалась также П. Гольдман-Ракич и ее коллегами на модели выработки отставленного инструментального глазодвигательного рефлекса у обезьяны. В этих опытах, так же как и в опытах С.А. Батуева, процедура отставленного ответа позволяла оценивать способность животного реагировать на те или иные ситуации, опираясь на представления о внешней среде, сохраняющиеся в рабочей памяти.
П. Гольдман-Ракич и ее коллеги рассматривают префронталь-ную кору как посредника между памятью и действием. В разных ее участках в режиме рабочей памяти кодируется информация о местоположении объектов, их признаках (цвете, размере, форме), а у людей — семантические и математические знания. Исследователи полагают, что префронтальная кора извлекает информацию из
долговременной памяти, гиппокамп же осуществляет консолидацию новых ассоциаций, которые так нужны для корректировки поведения с учетом недавно произошедших событий (см. раздел «Рабочая память»).
Существует и другое объяснение взаимосвязи обеих структур. Гиппокамп, так же как и префронтальная кора, причастен, к извлечению информации из долговременной памяти, которая ему необходима для сопоставления с текущими стимулами, чтобы вынести вердикт о новизне или его тождестве с прогнозом.
Сейчас уже многое известно о функциях моторной коры. Ее рассматривают как центральную структуру, управляющую самыми тонкими и точными произвольными движениями, посылающую свои сигналы к мотонейронам спинного мозга (так называемая пирамидная система). Именно в моторной коре строится конечный и конкретный вариант моторного управления движением. Моторная кора использует оба принципа управления; контроль через петли обратной сенсорной связи и через механизм программирования. Это достигается тем, что к ней сходятся сигналы от мышечной активности, от сенсомоторной, зрительной и других отделов коры, которые и используются для моторного контроля и коррекции движения.
Моторная кора включает первичную и дополнительную моторные области (ДМО), характеризующиеся соматотопической организацией с элементами множественного представительства периферии в этих отделах моторной коры- Первичная моторная кора расположена вдоль центральной борозды преимущественно в пре центральной извилине. В 5-м слое первичной моторной коры находятся гигантские клетки Беца, аксоны которых входят в состав пирамидного тракта, образующего эфферентные пути от моторной коры к мотонейронам спинного мозга. ДМО расположена в глубине межполушарной щели, примыкая к первичной моторной коре.
Клетки первичной моторной коры образуют колонки, которые возбуждают и тормозят группу функционально близких мотонейронов. Двигательная колонка представляет собой объединение нейронов, регулирующих работу нескольких мышц, действующих на сустав. При раздражении через микроэлектрод различных колонок возникают разнообразные движения в определенном суставе. Таким образом, в разных колонках представлены не отдельные мышцы, а разные движения. Это объясняет результаты А. Георго-пулоса (Сеог§ори1о5 А.) из университета Дж. Гопкинса, который нашел в моторной коре (в прецентральной извилине) обезьяны нейроны, кодирующие движения руки. Он вычислял векторы дви-
16-
жений на основе суммирования частоты импульсации многих нейронов, возбуждающихся перед движением. По его данным, вектор движения определяется результатом активности не более чем 100 нейронов. Он формируется за несколько миллисекунд до сокращения соответствующих мышц, приводящих руку в движение.
Премоторная кора (дорзальная и вентральная области) содержит представительство каждой ноги и руки, образуя прямые связи с мотонейронами спинного мозга. Дорзальная Премоторная кора, кроме того, имеет раздельное представительство дистальной и про-ксимальной части руки. Разряды нейронов дорзолатеральной пре-фронтальной и дорзальной премоторной коры тесно коррелируют со зрительно управляемыми движениями, характеризуясь селективностью в отношении направления движений конечностей.
Возникает вопрос: что именно отражает разряд нейронов в премоторной и префронтальной коре? Чтобы ответить на него, Г. Пеллегрино и С. Вайс (РеНееппо О., \У15е 5.) провели две серии опытов. В одной серии обезьяна двигала рычаг в направлении к цели, в другой — появление стимула в разных точках пространства служило сигналом к движению всегда в одном направлении. Стимул мог появляться в одной из 8 позиций при строгой фиксации взора на центр экрана. Оказалось, что премоторные нейроны разряжались избирательно по отношению к целевому движению, а не к самому движению как таковому. В отличие от них нейроны префронтальной коры отвечали избирательно на цель независимо от типа движения.
Важную функцию в управлении движением выполняет мозжечок. Он обеспечивает сохранение равновесия, поддержание позы, регуляцию и перераспределение мышечного тонуса, тонкую координацию движений. Нейроны моторной коры находятся под контролирующим влиянием мозжечка. В опытах с регистрацией нейронной активности у обезьян показано, что при выполнении ими заученного движения активность нейронов зубчатого ядра мозжечка на 10 мс опережает изменение активности нейрона в моторной коре, которое предшествует появлению мышечного движения. Влияние сигналов, поступающих из мозжечка, на активность нейронов моторной коры у обезьян также установлено в опытах с временным понижением температуры мозжечка. Во время его охлаждения импульсация нейронов моторной коры и соответствующее выученное движение запаздывали или вообще были невозможны.
Многие авторы отождествляют мозжечок с мощным процессором, в котором перерабатывается огромная информация. Полагают, что он обеспечивает временную избирательную настройку при выполнении любого двигательного акта, точное выполнение дви-
жения во времени. Поражение мозжечка ведет кдисметрии — плохому выполнению точных движений.
Кроме сигналов от мозжечка, в моторную кору поступают сигналы от базальных ганглиев — структуры, которая ответственна за хранение как двигательных программ врожденного поведения (пищевого, питьевого и др.), так и приобретенных навыков. Показано, что клетки базальных ганглиев, так же как и мозжечка, разряжаются задолго до движений, совершаемых животными в ответ на сигнал. По данным Н.Ф. Суворова (1980), разряды нейронов хвостатого ядра кошки на 50—150 мс опережают ЭМГ-ком-понент хватательного движения лапы, направленного на пищу, т.е. возникающего как натуральный пищевой рефлекс. В хвостатом ядре были найдены также нейроны, у которых возбуждение опережало условную двигательную пищевую и оборонительную реакции. Эти реакции у нейронов появлялись с выработкой условного рефлекса и исчезали с его угасанием. Нейроны с реакциями опережающими двигательные оборонительные и пищевые условные рефлексы, локализованы в различных частях хвостатого ядра.
Г. Могенсон и его коллеги (Мо§еп80пО.Ь.^опе50.1..,.лт С.]., 1980), рассматривая механизм локомоции и оральных двигательных актов, связанных с глотанием, поеданием пищи и питьем, в качестве ключевых структур мозга выделяют хвостатое ядро и прилегающее ядро стриатума (часть базальных ганглиев). Функция ХЯ — обеспечение произвольных действий в составе навыка. Прилегающее ядро (п.асситЬепз) имеет отношение к врожденному поведению. Инъекция ДА в прилегающее ядро вызывает у крыс локомоторные реакции в открытом поле- Электрическая стимуляция этого ядра вызывает жевательные и глотательные реакции. Прилегающее ядро находится под контролем вентрально-тегмен-тальной области (ВТО), которая получает ДА-ергические проекции от ствола мозга. Электрическим раздражением ВТО можно вызвать поведение атаки — нападения. ВТО является связующим звеном между лимбической системой и п.ассиптЬепк. Последнее имеет выход на бледный шар (рис. 53). Активация ВТО вызывает эффект «торможения торможения» (тормозных интернейронов), что обеспечивает ДА-ергическое возбуждение нейронов п.асситЬепя. Предполагают, что сигналы из ВТО к прилегающему ядру инициируют различные виды целенаправленного поведения (оборонительное, пищевое, питьевое).
Сходный механизм действует и в системе черная субстанция — ХЯ. Черная субстанция относительно ХЯ выполняет функцию, аналогичную ВТО для прилегающего ядра. Однако стимулами, запускающими эту цепь реакций, являются сигналы не из лимби-
| 1-3 0. и ^ ^ ЕС 0 Я ^ X 1^ Ю 3 г= о 5 я§ 0 У 0 <с | |||||||
| Черная субстанция | |||||||
| Е и ^ П '-" ^ 3§ |§ &§ 5 5 | |||||||
| ' | |||||||
| Хвостатое ядро (неостриатум) | |||||||
| & а '5 3 и | |||||||
| 0: и б « а з I ^ 1 ё 3 и 2: ^ | Прилегающее ядро (вентральный стриатум) | ^ я Н | |||||
| ———> | —»- | ||||||
| - | . | ||||||
| вто | |||||||
Рис. 53. Схема связей хвостатого ядра и прилегающего ядра — двух ключевых структур базальных ганглиев с корой и со структурами, распространяющими ДА-ергические влияния. Ассоциативная кора больших полушарий илимбическая система инициируют программы моторных актов через базальные ганглии. ВТО — вентрально-тегментальная область.
ческой системы, а из ассоциативной коры, и прежде всего из пре-фронтальной, которая имеет прямую проекцию на ХЯ. Через бледный шар ХЯ и прилегающее ядро запускают исполнение поведенческого акта, управляя моторными системами нижнего ствола мозга. Параллельно сигналы через таламус посылаются в моторную кору, которая включает контролирующий механизм, влияющий на выполнение действия.
Таким образом, моторная кора управляет движением, используя информацию, поступающую как по сенсорным путям от других отделов коры, так и от генерируемых в ЦНС моторных программ, которые актуализируются в базальных ганглиях и мозжечке и доходят до моторной коры через таламус и префронтальную кору.
Согласно гипотезе, высказанной П. Робертсом, актуализация моторных программ происходит вследствие активации командных нейронов. Сами командные нейроны могут контролироваться и затормаживаться сверху. Снятие торможения с командных нейронов повышает их возбудимость и тем самым высвобождает
«предпрограммированные» цепи для той деятельности, для которой они предназначены.
По-видимому, существует общий биохимический механизм актуализации моторных программ в результате роста активности ДА- и НА-ергических систем в мозге. Эти системы обладают свойством тормозить тормозные интернейроны и тем самым растормаживать нейронные сети.
Имеются данные о наличии в стриатуме ГАМК-ергических тормозных и нтер нейронов, которые тонически тормозят запрограммированные нейронные цепи, управляющие позой и движениями, а сами тормозятся ДА-ергическими нейронами из черной субстанции. Показано, что НА-волокна проецируются в мозжечок, их активация также ведет к торможению тормозных интернейронов.
Сейчас многие исследователи разделяют точку зрения, согласно которой ДА- и НА-ергические пути участвуют в подавлении активности тормозных интернейронов в структурах, вносящих вклад в инициацию и двигательный контроль поведения- Таким образом, в базальных ганглиях, мозжечке уже заложен механизм, который может актуализировать хранящиеся в них двигательные программы. Однако для приведения в действие всего механизма, необходимо, чтобы в эти структуры поступил сигнал, который послужил бы начальным толчком процесса.
|
|
|
|
|
Дата добавления: 2014-12-10; Просмотров: 564; Нарушение авторских прав?; Мы поможем в написании вашей работы!