КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Круг папецА 3 страница
|
|
|
|
ГЛАВА III
Возможно также, что импульсы, возникающие в рецепторах различных сенсорных модальностей, как висцеральных, так и соматических, поступают в лимбические цепи через диффузные восходящие пути. Известно, что различные специфические афферентные волокна дают многочисленные коллатерали к ретикулярной формации. Кроме того, существуют афферентные системы, подходящие к этой цепи от таламуса и базальных ганглиев, а также от коры, и эфферентные системы, через которые лимбические структуры могут влиять на активность таламуса и коры. Пути, по которым идут разряды от гипоталамуса и лимбических областей среднего мозга, показаны на фиг. 5 пунктирными линиями, направляющимися к ретикулярной формации ствола. Последняя передает эти сигналы как вверх, так и вниз, воздействуя таким образом на центры спинного и головного мозга.
ЛИМБИЧЕСКАЯ ДОЛЯ И РАЗВИТИЕ ПРИМИТИВНОГО МОЗГА
В сравнительном исследовании мозга млекопитающих, опубликованном в 1878 г., Брока [104] отметил стабильность кортикальных структур, граничащих со стволом мозга у входа в полушария. Он приписывает честь открытия этой «доли» Герди и Фовиллу, описавшим ее за 40 лет до него. Однако использованные ими термины вышли из употребления, а общепринятым является описательный термин Брока «большая лимбическая доля». Новая кора и лежащее под нею белое вещество, состоящее из нервных волокон, соединяющих кору с более древними корковыми и подкорковыми структурами, проходят грандиозный путь развития в ходе эволюции, но лимбические структуры остаются относительно неизменными. Однако мощное разрастание новых структур приводит к изменениям и перемещениям, так что некоторые старые связи трудно проследить. В последующих разделах мы попытаемся указать на некоторые наиболее важные из этих изменений.
Мозг развивается из передней части нервной трубки, где ее выпячивания образуют боковые желудочки полушарий. До появления этих выпячиваний передний
НЕРВНЫЙ СУБСТРАТ ЭМОЦИЙ
мозг эмбриона млекопитающего на ранней стадии развития имеет в основном цилиндрическую форму, причем
стенка состоит из массы [553]. Как показывает фиг. 6, эти массы клеток являются зачатками гип-покампа, перегородки, полосатого тела, пириформ-ной коры и неокортекса. На более поздних стадиях развития в связи с разрастанием коры и лежащих под ней волокон (в том числе массивного мозолистого тела) полосатое тело оттесняется в подкорковую область, а другие структуры первичной коры (гиппокамп, перегородка и пириформная кора) смещаются к центру. Новая кора и белое вещество полушарий буквально подавляют эти первичные корковые структуры.
Ринэицефалон, или первичный обонятельный мозг, занимает сравнительно небольшую часть полушарий мозга высших млекопитающих. Слух и зрение, которые дают гораздо более обширную информацию об окружающем мире, чем обоняние, становятся более важными для направления поведения организма в целом. Новая кора с огромным количеством нервных клеток и связей развивается
плотноупакованных клеток

Фиг. 6. Основные отделы коры головного мозга человека на разрезе [5S3].
А. У эмбриона 15 мм. Б. У эмбриона
50 мм. В. У взрослого (основная массч
гнпнокампа у взрослого сдвинута назал..
в плоскость разреза но попала).
ГЛАВА III
в основном в связи со слуховыми и зрительными дистантными рецепторами, особенно в связи с органом зрения и возникновением исполнительных механизмов, посредством которых можно влиять на зрительную информацию.
Однако первичная кора (с ее подкорковыми придатками) не исчезает и не утрачивает своих функций. Следует признать, что ее функции не вполне ясны и в какой-то части гипотетичны, но несомненно одно, что эта кора играет важную роль в регуляции висцеральных функций и эмоциональных процессов, связанных с внутренними состояниями. Мак-Лин [600] назвал эту кору «висцеральным мозгом», а последние исследования пролили свет на ее роль в организации эмоций и мотивации поведения. После описания основных анатомических связей мы рассмотрим экспериментальные данные, которые подтверждают важную роль лимбических и гипоталами-ческих структур в эмоциональных процессах.
Афферентный обонятельный путь начинается биполярными нейронами в обонятельном эпителии. Дисталь-ные концы этих нейронов прободают кутикулу эпителия и служат обонятельными рецепторами. Тела нейронов лежат среди опорных клеток обонятельного эпителия, а их короткие аксоны идут в составе обонятельного нерва через горизонтальную пластинку решетчатой кости, оканчиваясь в обонятельной луковице. Последняя.представляет собой первичный обонятельный центр [605]. У низших млекопитающих и у эмбрионов высших млекопитающих на ранних стадиях развития обонятельная луковица — это не что иное, как наиболее ростральная часть первой, или обонятельной, доли полушарий. В мозге человека она представляет собой относительно небольшое образование, лежащее на основании мозга; от этого образования берет начало обонятельный тракт, который в каудальном направлении делится на латеральную, медиальную и промежуточную полоски. Промежуточная полоска сразу же погружается в переднее продырявленное вещество (или у некоторых видов — в обонятельные бугорки). У человека она мала, непостоянна и, вероятно, не имеет большого функционального значения.
Чтобы понять анатомические связи обонятельных структур ствола мозга и новой коры, необходимо знать
НЕРВНЫЙ СУБСТРАТ ЭМОЦИИ
их эволюцию. На фиг. 7 схематически показано развитие медиальной обонятельной полоски. В мозге амфибии (тритона) обонятельная луковица — это фактически обонятельная доля, наиболее ростральная часть головного мозга. Волокна, идущие от нее каудально, по направлению к медиальным частям переднего мозга (гиппо-
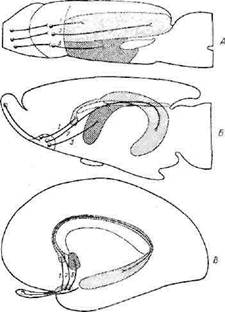
Фиг. 7. Схема эволюции медиальной обонятельной полоски [553].
Схемы связей у тритона (А), крысы (Б) и человека (В). Переднее обонятельное ядро связано с передним отделом гмппокампа (/). с задним отделом гиппокампа (2) и с ядрами перегородки (3). У человека длинный рудимент гипгюкампа располагается нал мозолистым телом.
ГЛАВА III
камп и перегородка) заканчиваются синапсами на нейронах переднего обонятельного ядра.
Переднее обонятельное ядро у высших животных состоит из клеток, переместившихся из обонятельных луковиц через тракт и полоски на основание полушарий. На схемах мозга крысы и человека для простоты оно изображено компактным. Только часть волокон тракта заканчивается в этом ядре; остальные, не прерываясь, идут далее, ко вторичным центрам.
Следует отметить, что большая часть гйппокампа смещена у крысы в вентральном и каудальном направлениях. У человека перемещение гйппокампа продолжалось, пока он не занял своего положения в височной доле, оставив лишь небольшой «рудимент», над мозолистым телом (см. фиг. 7). Этот рудимент указывает направление перемещения собственно гйппокампа и поддерживает связь между ним и структурами в области перегородки. Не ясно, существует ли прямая нервная связь между обонятельной луковицей и гиппокампом [725]. Для проводимого нами анализа не существенно, имеют ли чисто обонятельные по функции волокна, идущие от обонятельных луковиц, прямые связи с отдельными рннэнцефальными структурами или подходят к ним только после синаптического переключения.
Латеральная полоска идет к структурам,, лежащим на основании полушария — к пириформной области — и миндалевидному ядру. Глур [382] рассматривает миндалевидное ядро как подкорковое скопление серого вещества, принадлежащее к обонятельному мозгу, или лим-бической системе. Среди его афферентных связей четко определены анатомически лишь обонятельные волокна, достигающие его по латеральной обонятельной полоске [382]. Поэтому мы можем рассматривать миндалевидное ядро как вторичный обонятельный центр, подобно другим структурам, к которым подходят вторичные обонятельные волокна от обонятельной луковицы и переднего обонятельного Ядра. (Это довольно широкие обобщения, но они полезны для того, чтобы получить общее представление об обонятельном мозге.)
Эллиот [220] представил простую и удобную схему, иллюстрирующую эволюцию некоторых основных обо-
НЕРВНЫЙ СУБСТРАТ ЭМОЦИИ
нятельных путей и подчеркивающую важные связи, существующие между ними и гипоталамусом (фиг. 8). Первичные обонятельные нейроны (рецепторы) связаны

Фиг. 8. Схема эволюции основных обонятельных путей [220].
Среднесагиттальный разрез мозга лягушки (А) и человека (£). Первичные и вторичные пути (тонкие линии) связывают обонятельный эпителий с обонятельной луковицей и полушарием, / — обонятельный бугорок; 2 —перегородка; S — гиппокамп; 4 — гипоталамус. Из полушария пути после переключения идут по длинной дуге в гипоталамус. Перегородка и обонятельный бугорок связаны с гнппокампом более прямым путем, медиальным передне-мозговым пучком (свод).
с первичным обонятельным центром, т. е. с обонятельной луковицей. Вторичные нейроны, лежащие в луковице, непосредственно связаны с 1) обонятельным бугорком (переднее продырявленное вещество), 2) перегородкой, 3) гнппокампом и 4) гипоталамусом. (Переднее обонятельное ядро, в котором происходит переключение
ГЛАВА III
многих из этих волокон, а также волокна латеральной полоски здесь не изображены.) Мы считаем нужным особо подчеркнуть тот факт, что гипоталамус получает прямые волокна не только от обонятельной луковицы, но и от связанных с ней переключающих ядер. Один из этих постсинаптических путей, а именно путь, идущий от гиппокампа, образует дугу, располагаясь над верхним концом ствола мозга (отсюда его название свод). Другие волокна, идущие от вторичных обонятельных центров (бугорок, или переднее продырявленное вещество, и перегородка), а также волокна от первичного центра (обонятельная луковица) идут по довольно прямому пути к гипоталамусу в составе медиального передиемозго-вого пучка. Последний представляет собой древний путь, который благодаря своему положению у основания мозга почти не изменяется в ходе эволюции. Этот комплекс волокон связывает вентро-медиальные области полушарий мозга с преоптическои и гипоталамическои областями [522]. Ларселл [562] утверждает, что в его составе есть восходящие и нисходящие волокна, проходящие вдоль всей латеральной гипоталамическои области, отдающие волокна к ядрам гипотоламуса и (по данным некоторых исследователей) распространяющиеся до покрышки среднего мозга. Эти связи можно видеть на фиг. 9.
Наута [679] описывает медиальный передпЪчозговой пучок как большой древний путь, содержащий значительную часть связей, соединяющих лимбическую кору с гипоталамусом, центральным серым веществом и ретикулярной формацией среднего мозга. На фиг. 9 выделен медиальный переднемозговой пучок, но изображены только его восходящие волокна. Можно видеть, что значительное число волокон идет от ядер покрышки среднего мозга и центрального серого вещества через гипоталамус. Рострально от гипоталамуса эти волокна поворачивают к перегородке и миндалевидному ядру, которые, таким образом, очевидно, являются основными областями конвергенции афферентных обонятельных импульсов и восходящих импульсов от структур ствола мозга. Фиг. 9 представляет собой полезное схематическое обобщенное изображение основных элементов лим-
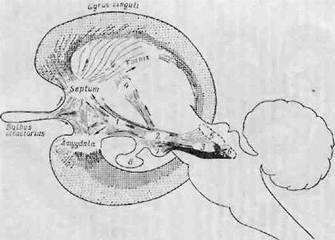
Фиг. 9. Схема лимбической доли [604].
Архикортекс показан штриховкой, а палеокортекс—пунктиром. Выделен медиальный пе-реднемозговой пучок I — основная связь между лимбической долей, гипоталамусом {2) и лимбической областью (3), которая состоит из центрального серого вещества и околоцевт-ральной ретикулярной формации среднего мозга и осуществляет жизненно важную связь между лимбической корой, нижней частью ствола мозга и спинным мозгом; 4 — переднее ядро таламуса; 0 — центральное серое вещество среднего мозга, в — вентральное и дорсальное ядра покрышки; 7 — маммиляр-ное тело; 8 — гипофиз.
ГЛАВА III
бпческой системы. Примитивная древняя кора (архикортекс), особенно гиппокамп и его рудимент (связанный с поясным пучком), окружена филогенетически более поздней старой корой (палеокортекс), состоящей в основном из поясной и гиппокамповой извилин. Архикортекс и палеокортекс с их афферентными и афферентными связями, функционируя вместе с гипоталамусом и ретикулярной формацией ствола мозга, регулируют висцеральную активность организма и обеспечивают «внешнее выражение внутренних состояний». Однако эмоции в нашем понимании, в том числе аффект и его выражение, требуют для своего осуществления взаимодействия между этими структурами и новой корой (неокортекс). Только последняя с ее пластичностью, способностью сопоставлять информацию различных' видов, хранить информацию и сравнивать явления, происходящие в данный момент, с прошлым опытом,— только такая структура может обеспечить осмысленный опыт и сознание. Таким образом, необходимо учитывать анатомические связи, которые, очевидно, представляют собою цепи, сформировавшиеся специально для обеспечения указанных взаимодействий гипоталамуса, лимбической системы и новой коры.
Папец [692] в 1937 г. писал: «Эмоции — это настолько важная функция, что, каков бы ни был их механизм, он должен иметь морфологическую основу». Он предположил, что такой морфологической основой являются гипоталамус, поясная извилина, гиппокамп и их связи. Мак-Лин [600] представил наглядную схему (фиг. 10), иллюстрирующую представления Папеца. Папец указывал, что в мозге низших позвоночных медиальная стенка полушария анатомически и функционально связана с гипоталамусом, а латеральная стенка — с дорсальным таламусом [692]. По мнению Папеца, структуры медиальной стенки полушария со временем превращаются в гиппокампову формацию, которая посылает импульсы через свод к гипоталамусу (Маммилярные тела, см. фиг. 9). Таким образом, свод становится первой эффе-
НЕРВНЫЙ СУБСТРАТ ЭМОЦИИ
рентной связью коры (архикортекса) с гипоталамусом. Позднее с появлением поясной извилины (палеокортекс) ее афферентные волокна связывают маммилярные тела с передним ядром таламуса, а уже оттуда в составе
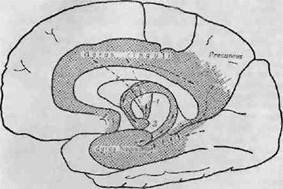
Фиг. 10. Круг Папеца [600а].
Заштрихованная область коры представляет лимбическую долю Брока, или
висцеральный мозг Мак-Лина. / — переднее ядро таламуса; 2—маммилярцое
тело.
проекционных волокон таламуса подходят к этой извилине. Далее, Папец предположил, что от поясной извилины импульсы могут идти назад к гиппокампу через поясные пучки. Он высказал предположение, что «гипоталамус, передние ядра таламуса, поясная извилина, гиппокамп и их взаимосвязи составляют гармонически работающий механизм, лежащий в основе возникновения эмоций и участвующий в их выражении». Это предположение основано на полученных в эксперименте и клинике данных об эмоциональных расстройствах, возникающих в результате повреждений какого-либо из звеньев этого круга. Папец предполагал, что эмоции возникают либо сначала в коре, причем импульсы всту-
4—517
ГЛАВА III
лают в круг через гиипоками, либо в результате возбуждения гипоталамуса. В последнем случае «кору поясной извилины следует рассматривать как воспринимающую область для эмоциональных ощущений, возникающих в результате прихода импульсов из области гипоталамуса, по аналогии со стриарной областью, которая воспринимает зрительное возбуждение, идущее от сетчатки» [692].
Вероятно, эмоция, возникшая любым путем, приводит к генерализованной активации круга Папеца. Палец достаточно осторожно указал, что «к гипоталамусу могут идти как висцеральные, так и соматические импульсы из многих периферических источников», которые, как хорошо известно, придают эмоциональную окраску сенсорным восприятиям. Он предположил, что афферентные импульсы от различных рецепторов на уровне тала-муса делятся на три «потока»: «поток движения», «поток мысли» и «поток чувства». Первый проводит импульсы через дорсальный таламус к полосатому телу, второй — от таламуса к латеральной коре, а третий — через вентральный таламус к гипоталамусу, а оттуда через маммило-таламический тракт к переднему ядру таламуса, которое связано с поясной извилиной. Одновременная активность второго и третьего потоков должна обеспечивать сенсорный опыт (второй поток) и эмоциональную окраску (третий поток).
В качестве убедительного доказательства того, что гиппокамп входит в состав эмоционального круга, Па-пец привел интересный факт: при бешенстве, ведущим симптомом которого являются интенсивные эмоциональные (реакции, специфическое поражение локализуется главным образом в гиппокампе и супраоптическом ядре.
Однако в целом данные, иа основе которых Папец создал свою теорию механизма эмоции, были весьма скудны, и ему приходилось делать далеко идущие выводы из того, что было известно в то время. Мак-Лин [600], рассматривая «висцеральный мозг» в связи с психосоматическими заболеваниями, указал в 1949 г., что многочисленные экспериментальные исследования, проведенные со времени опубликования статьи Папеца,
нервный субстрат эмоции
подтвердили представления последнего о первостепенной роли обонятельного мозга в аффективной сфере.
Теоретическая статья Папеца [692] была одной из двух ранних попыток определить механизмы эмоций. Другой попыткой было сообщение Клювера и Бью си [545] о резких изменениях эмоционального поведения макакрв-резусов при обширных повреждениях височной доли. Эти две статьи появились с интервалом всего в несколько месяцев; интересно, что Брэйди [90] в своем недавнем обзоре, посвященном проблеме «эмоционального поведения», подчеркнул, что данные, накопленные со времени опубликования этих статей, предоставляют в наше распоряжение богатые анатомические, физиологические и поведенческие доказательства удивительной прозорливости этих исследователей.
Хотя гипотеза Папеца хорошо продумана и служит прочным фундаментом для современных представлений о механизмах эмоций, она имеет один весьма существенный недостаток: она игнорирует несомненно важное участие новой коры в этом механизме. Остается еще много выяснить относительно функциональных связей между новой корой и гипоталамо-лимбической системой; несомненно, однако, что такие связи существуют и весьма многочисленны. Глур [381] предположил, что старые образования переднего мозга (в том числе гиппо-камп, гиппокампова извилина, поясная извилина, миндалевидное ядро и бледный шар) «действуют как посредники между филогенетически более молодым конечным мозгом и более древними частями промежуточного мозга — гипоталамусом и системой уздечки (эпитала-мус), с которыми эти «новые» области, за исключением поля 6, не имеют более прямых связей».
Следует еще раз подчеркнуть стратегическое положение гипоталамуса, поскольку он связывает лимбиче-ские структуры переднего мозга с лимбической областью среднего мозга (обеспечивая таким образом жизненно важную связь с нижним отделом ствола и спинным мозгом), а также поскольку он обладает существенными реципрокными связями с корой. Наконец, необходимо помнить следующее: 1) в основе поведения лежат висцеральные функции, а гипоталамус и архипал-
4*
100 ГЛАВА III
лиум — рано возникающие в процессе эволюции — осуществляют регуляцию этих функций; 2) выражение внутренних состояний становится возможным с развитием промежуточной системы — «выроста» внутренней системы, соединяющей последнюю со скелетными мышцами через ряд подкорковых ядер, и 3) сознательное восприятие и произвольная регуляция, возникающие при дальнейшем разрастании центральных структур, сохраняют тесную связь с этими более ранними по происхождению системами.
Физиолог и психолог должны установить, каким образом при взаимодействии этих тесно связанных систем интегрируются все функции организма, в том числе аффекты и выражение эмоций.
ГЛАВА
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ
эмоций
ВЫРАЖЕНИЕ ЭМОЦИИ И ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
Объем книги не позволяет нам дать полный обзор многочисленных клинических и экспериментальных данных, подтверждающих наше убеждение, что гипотала- I мо-лимбическая система имеет важнейшее значение для эмоциональных ощущений и выражения эмоций. Одной из самых узловых проблем в этой области являются поиски надежных индикаторов эмоциональных состояний. Монументальный труд Кэниона [133] представляет собой классическую попытку в этом направлении (см. также [204]). Однако выбор надежных критериев для определения эмоционального состояния все еще представляет большие трудности, если учесть тот факт, что большинство реакций, связанных с эмоциями, может возникать, и действительно возникает, также и в отсутствие эмоций [90]. Попытки сопоставить физические сдвиги с эмоциями в основном базировались на периферических реакциях, таких, как изменение кожной температуры, потоотделение, пилоэрекция, изменения секреторной активности желез, кровяного давления, частоты сердечных сокращений и т. д.
Общепризнано, что многие соматические реакции служат индикаторами эмоций. Выражение лица, съеживание, выгибание спины, ожесточенный бег по кругу, нападение или бегство — вот несколько хорошо известных примеров. Мы считаем эти поведенческие реакции непроизвольными, хотя они опосредуются соматической нервной системой и в различной степени подчиняются произвольной регуляции. Внутренние органы почти совсем не подвластны воде, и общепризнано, что их ре-
ГЛАВА IV
акции, т. е. изменения, регулируемые вегетативной нервной системой, должны быть, следовательно, более надежными показателями эмоциональных состояний, нежели «соматические» реакции. Однако и в этом случае вероятность ошибки довольно велика, так как основная функция вегетативной регуляции заключается в обеспечении гомеостаза. При слабо выраженных изменениях эмоционального состояния или настроения соответствующие физикальные сдвиги могут быть столь незначительными и охватывать столь различные органы, что по ним очень трудно установить определенные «психосоматические» взаимоотношения.
Рассмотрим, например, изменения сердечного ритма. Кардиальные условные рефлексы вырабатываются довольно легко [201], нередко после первого знакомства индивидуума с новой ситуацией, и, возникнув однажды, могут сохраняться в течение нескольких лет без подкрепления. В таком случае кардиальный условный рефлекс можно вызвать у индивидуума, внешне спокойного, но реагирующего эмоционально на ситуационные раздражители, которые на основании других критериев можно отнести к подпороговым- Таким образом, частота сердечных сокращений может служить чувствительным критерием эмоций, но, к сожалению, она столь же тонко реагирует и на многие другие сдвиги.
Ввиду отсутствия какого-то одного надежного показателя эмоциональных процессов многие исследователи пытались подобрать группу индикаторов, которые служили бы таким критерием в сочетании. Например, предпринимались попытки разработать специальную «шкалу», которая позволила бы количественно оценивать сдвиги в разнообразных реакциях, отражающих эмоциональное состояние организма, возникающее при повреждении лимбической системы [89]. Самое сложное в подобных случаях — это дать количественную оценку самой эмоции. Всякое научное объяснение заключается в отыскании постоянных связей между явлениями, и любой тест, применяемый при научном исследовании, состоит в том, что одна из переменных измеряется косвенно, посредством измерения другой переменной, являющейся функцией первой. Разработка такого теста предусматри-
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ ЮЗ
вает независимое изменение обеих переменных до тех пор, пока не будет выведен закон их связи [516]. Итак, проблема сводится к тому, чтобы дать количественную оценку эмоции при одновременном измерении различных сопровождающих ее телесных реакций. Очевидно, нам приходится довольствоваться той субъективной оценкой, какую дает своим эмоциям сам испытывающий их человек. Несомненно, это связано с серьезными трудностями и ошибками. Однако такова наша отправная точка, и из этого мы должны исходить, пользуясь менее прямыми (но более объективными) методами, которые убеждают нас в том, что такие внешние проявления, как выражение лица, движение хвостом, различные звуки, изменения кровяного давления и частоты сердечных сокращений, пилоэрекция, кожное сопротивление и т. д., указывают на определенные виды и степени эмоциональных реакций. Несомненно, что выявление качественных индикаторов — не столь трудная проблема, как установление точных количественных критериев.
Хотя эмоция представляет собой сложное и, казалось бы, неуловимое явление, существующие клинические и экспериментальные данные показывают, что мы можем судить об эмоциональных ощущениях, наблюдая за выражением эмоций. Мы считаем вполне убедительными доказательства, подтверждающие предположение многих авторов о том, что лимбо-гипоталамический комплекс обладает первостепенной важностью, по крайней мере как основная часть нервного субстрата эмоции [87, 177, 179, 267, 520, 543, 544, 546, 580, 602, 692, 725, 848, 911, 944].
ПОВРЕЖДЕНИЯ, ПРИВОДЯЩИЕ К ПОЯВЛЕНИЮ РЕАКЦИИ ЯРОСТИ
Одно из наиболее ярких доказательств участия ги-поталамо-лимбической системы в организации эмоций обнаруживается при повреждениях мозга, которые превращают ручных животных в диких зверей. Уже в 1884 г. было отмечено, что у декортицированных собак очень легко вызвать ярость [389]. Это было многократно
ГЛАВА IV
подтверждено в опытах на собаках и кошках [35, 38, 136, 209, 773], но само по себе еще не доказывало роли гипоталамуса или лимбической системы.
Удаляя различные части мозга у кошек, Бард [35] показал, что каудальная область гипоталамуса имеет особое значение для полного проявления реакции ярости, возникающей у декортицированных животных. Однако позднее было обнаружено, что хотя каудальная область гипоталамуса необходима для полного проявления реакции ярости, ограниченные реакции такого рода возникают у кошек-и при «высокой» и «низкой» перерезке среднего мозга [41]. В 1925 г. Кэннон и Бриттон [136] ввели термин «мнимая ярость» для обозначения этого поведения, очень напоминающего реакцию ярости у интактного животного. Бард [35] принял этот термин, но позднее разъяснил, что эпитет мнимая вовсе не означает, что речь идет о какой-то «псевдоярости»; он использован лишь для того, чтобы исключить какие бы то ни было рассуждения об эмоциональных ощущениях у животного, лишенного коры [37]. Он утверждал, что такое поведение (мнимая ярость), несомненно, аналогично реакции ярости, наблюдаемой у нормальной кошки. Можно сделать вывод, что у интактного бодрствующего животного такое поведение сопровождается эмоциональным ощущением, и единственным прочным основанием для такого вывода является признание того факта, что мы сами испытываем такие ощущения параллельно с указанным проявлением.
Бард показал также, что мнимая ярость возникает у декортицированных кошек при тех же условиях, которые вызывают отчетливые реакции ярости у нормальных животных, и что так же, как и у нормальной кошки, ее легко отличить от других форм эмоциональных проявлений, таких, как страх или половое поведение. Поскольку вызвать реакцию мнимой ярости у декорти-цированного животного очень легко, Бард [38] полагает, что в норме кора каким-то образом тормозит механизм интеграции этой реакции, короче говоря, что каудальная область гипоталамуса обычно тормозится новой корой, а при декортикации она освобождается от этого влияния. Позднее на основе опытов с более огра-
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ
ничейными разрушениями он пересмотрел свое мнение [40].
фултон и Ингрэм [269] производили двухсторонние разрушения основания лобных долей как раз над хиаз-мой и обнаружили, что у кошек возникает реакция мнимой ярости. К сожалению, не удалось установить, какие важные для реакции пути перерезаются при этом. Фултон [266] позднее утверждал, что у кошки или обезьяны можно удалить оба лобных полюса и реакция ярости при этом не возникает; поэтому «вряд ли вероятно, что эффект был связан с путями, идущими от коры». Отметив, что большинство исследователей экспериментально вызываемого одичания, наносили очень обширные повреждения, Уитли и др. [502, 943] предприняли успешную попытку найти небольшую, четко определяемую область, разрушение которой приведет к указанному эффекту. Такой областью оказалось вентро-медиальное ядро гипоталамуса, при разрушении которого дружелюбное животное становилось исключительно злобным. (Попутно они отметили у таких животных гиперфагию и ожирение, что было подтверждено рядом позднейших исследователей, изучавших механизмы голода.) Описан интересный клинический случай: у больного, у которого при жизни отмечались периоды возбуждения, граничащие с яростью, после смерти было обнаружено поражение большинства медиальных ядер гипоталамуса [19]. В другом клиническом случае у девушки, склонной к приступам ярости, в области соединения миндалевидного ядра и гиппокампа была обнаружена опухоль величиной с вишню [910].
Противоречивые данные были получены при удалении орбитальной области: одни исследователи, производившие эту операцию у кошек, наблюдали мнимую ярость [527], а другие, наносившие повреждение в области поля 13 у обезьян, описали противоположный эффект— животное становилось ручным [589]. При повторных исследованиях было установлено [40], что удаление одной только коры не сопровождается реакциями ярости, хотя прежде Бярд [38] считал, что эти реакции связаны с освобождением гипоталамуса из-под тормозных влияний коры. По-видимому, склонность к реакци-
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 720; Нарушение авторских прав?; Мы поможем в написании вашей работы!