КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Разрушения, делающие животных более ручными 3 страница
|
|
|
|
Это объяснение приложимо равным образом и,к парасимпатической системе, поскольку у разных индивидуумов в состоянии эмоционального возбуждения наблюдаются выраженные количественные различия в ее реактивности. Обморочное состояние вследствие резкого падения кровяного давления, сопровождающегося замедлением сердечного ритма, часто возникает при виде крови или в состоянии страха, когда человеку что-то
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 145
угрожает. Симптомы повышенной реактивности парасимпатической системы часто наблюдаются у невротиков и лип с навязчивыми состояниями [895].
Приведенные выше примеры показывают, что типы эмоциональных вегетативных реакций тесно 'связаны с особенностями личности. Если бы мы научились изменять вегетативную реактивность на уровне гипоталаму-j са, это «южно было 'бы использовать для лечения невро-1 зов и психозов. Прежде чем проводить дальнейший анализ этих проблем, мы на1мерены обсудить исследования, иллюстрирующие возможность изменения реактивности гипоталамуса у экспериментальных животных. Если мы примем, что в основе ощущения и выражения эмоций лежат гипоталамические разряды, то мы тем самым признаем, что открытие путей регуляции реакций гипоталамуса равносильно обнаружению способов регуляции самих эмоций. Потенциальное значение такой регуляции для психического (да и физического) здоровья со всеми его последствиями для личности и для общества трудно переоценить. Мы считаем, что весьма важные первые шаги к пониманию регуляции реактивности гипоталамуса и связанных с нею эмоциональных реакций уже сделаны, и мы обсудим их ниже. Экспериментальные данные,.которые мы опишем, не дают полного ответа на вопрос о путях регуляции гипоталамуса — до этого еще далеко. Мы можем только надеяться, что эти данные окажутся направляющими для дальнейшей исследовательской работы и укажут пути ее плодотворного развития.
ПОНЯТИЕ «НАСТРОЙКИ» В НЕРВНОЙ СИСТЕМЕ
Из повседневного опыта известно, что реакция человека на данную ситуацию в значительной степени зависит от его умственного, физического и эмоционального состояния. Можно сказать, что человек «настроен» реагировать определенным образом. Вегетативные реакции на данный раздражитель могут быть то в основном симпатическими, то в основном парасимпатическими. Можно сказать,, что вегетативная система «настроена» таким образом, что тот или иной ее отдел оказывается макси-
146 /ГЛАВА V
мально чувствительным. Раздражитель, обладающий способностью возбуждать оба отдела, может вызывать реакции, различающиеся^ степени участия в «их.каждого из этих центров. Сенсибилизация вегетативных центров была обозначена термином настройка [310], так что принято говорить о симпатической или парасимпатической настройке. В наш век электроники термин настройка обычно ассоциируется с настройкой приборов на резонансную частоту. Поэтому мы должны подчеркнуть, что в этих рассуждениях этот термин не имеет отношения к частоте и указывает только на сенсибилизацию или облегчение определенных центров головного (и спинного) мозга.
Настройка в соматической нервной системе
Рефлекторные движения, такие, «ак сгибание задней конечности в ответ на укол в область подошвы, как правило, легко воспроизводятся. Качественно и количественно ответ остается постоянным, если то же раздражение наносится с достаточными интервалами. Это можно наблюдать на фоне глубокого наркоза, при котором высшие отделы центральной нервной системы невозбудимы, или на спинальном животном, у которого головной и 'спинной мозг разобщены. Это так называемые спинальные рефлексы, которые свободны от каких бы то ни было влияний со стороны головного мозга, что делает их значительно более устойчивыми и надежными. Благодаря этому спинальное животное представляет собой идеальный препарат для экспериментального изучения способов воздействия на рефлексы, осуществляемые помимо головного мозга.
Экспериментальное воздействие на спинальные рефлексы легче.всего продемонстрировать при раздражении одной конечности (укол, электрическое раздражение или постукивание по сухожилию) и наблюдении за реакцией другой конечности. Даже при повторении одного и того же раздражения и постоянстве ответа одной конечности реакция другой вырьирует в широких пределах в зависимости от ее исходного положения. Можно показать, что при раздражении одной задней конечности
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 147
другая либо сгибается, либо разгибается (в бедренном, колейном или голеностопном суставе) в зависимости от того, в катом 'положении она.находилась до.раздражения. Если она 'была согнута, то произойдет разгибание; если она была разогнута, то раздражение приведет к сгибанию [615а].
Каков механизм, который изменяет реактивность нервной системы спиналыного животного таким образом, что рефлекторный ответ качественно меняется? Эксперименты показали, что после.перерезки афферентных связей конечности со спинным мозгом ее исходное положение не влияет на ее ответ при раздражении противоположной конечности. Теперь нам известно, что эти нервы передают спинному мозгу сигналы, которые информируют его о длине и степени напряжения мышц. Достигая головного мозга, эта информация играет существенную роль в восприятии.положения конечности. Благодаря этому чувству положения, передающемуся через проприорецепторы и кожные рецепторы, даже слепой человек имеет точное представление о расположении частей своего тела.
Хотя опиналь/ное животное или животное, находящееся в состоянии наркоза, не воспринимает положения своих конечностей, это положение тем ее менее играет важную роль в определении характера рефлекторного ответа. Описанное выше влияние позы на рефлекторный ответ и исчезновение этого влияния после перерезки афферентных корешков свидетельствуют об определяющей роли афферентных сигналов от «рецепторов позы» в раздражении мотонейронов. Среди этих рецепторов особое значение для наших рассуждений имеют проприорецепторы самих мышц, мышечные веретена. Очевидно, они оказывают огромное влияние и могут менять рефлекторные реакции даже после устранения рецепторов сухожилий и суставов [615а].
Импульсы проприорсцепторов, изменяющие возбудимость опиналыных мотонейронов, создают различные виды «настройки». При контралатеральных рефлекторных реакциях, описанных выше, мы можем говорить о «настройке сгибателей» или «настройке разгибателей» в зависимости от того, каких мотонейронов касается
148 ГЛАВА V
облегчение, определяющее характер ответа,— мотонейронов сгибателей или разгибателей.
Известно далее, что такая настройка.нервной системы 'может быть достигнута совершенно иным способом. Вместо изменения притока импульсов к спинному мозгу путем изменения положения конечности можно изменять реактивность самого спинного мозга. Это происходит, например, три утомлении центров вследствие чрезмерного раздражения,.при шоке в результате тяжелого кровотечения. При этих условиях раздражитель, который прежде вызывал рефлекс сгибания, вызывает разгибание [306].
Интересно отметить, что явления настройки не ограничиваются спинным мозгом, они касаются и высших уровней центральной нервной системы. Так, движения, вызываемые электрическим раздражением моторной зоны коры головного мозга, также могут меняться в зависимости от условий эксперимента [363, 364].
Первые данные о настройке в вегетативной нервной системе
Аналогичные наблюдения были сделаны и в отношении вегетативной нервной системы. Особое значение для рассматриваемой нами проблемы имеет то, что такие наблюдения были проведены на человеке [330, 352]. Известно, что при произвольном сокращении группы мышц кровоток через них возрастает. Это несомненно полезная реакция, поскольку она способствует обеспечению возросшей потребности мышцы в кислороде и освобождению ее от образующихся в избытке продуктов обмена. Однако усиление кровотока не ограничивается работающими мышцами, а распространяется и на соответствующие мышцы на другой стороне. Это позволяет предполагать, что изменение кровотока обусловлено изменением активности центральной нервной системы, а не является простым следствием локального сосудорасширяющего влияния продуктов обмена на кровеносные сосуды сокращающейся мышцы. Это предположение подтверждается тем, что при произвольном сокращении усиление кровотока через мышцы несколько предшествует
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 149
самому сокращению. Создается впечатление, что процесс подготовки нервной системы к произвольному движению предусматривает такое изменение активности сердечно-сосудистых центров, которое обеспечивает усиление 'притока питательных веществ туда, где они будут необходимы.
Еще более поразительны результаты опытов по сравнению кровотока в утомленной и неутомленной мышце. Испытуемые выполняли произвольные мышечные движения. В утомленных мышцах вместо повышения кровотока наблюдается реакция (Противоположного характера. В связи с этим возникает вопрос, посылает ли центральная нервная система в обоих случаях одну и ту же информацию, а противоположный характер реакции (снижение кровотока) зависит от состояния, в котором находится утомленная мышца, или изменение реакции зависит от изменения настройки центрального механизма в состоянии утомления. Два наблюдения поддерживают последнее предположение: 1).произвольное сокращение утомленной группы мышц ведет к снижению кровотока не только через них, но также и через соответствующие (неутомленные) мышцы противоположной стороны; 2) после утомления мышц одной руки сокращение соответствующей неутомленной группы мышц (Противоположной руки приводит к повышению кровотока в обеих группах [353, 930]. Ясно, что в этих условиях повышение или понижение кровотока определяется не состоянием самих мышц. Если (Повышение кровотока диктуется мышце центральными механизмами, кровоток возрастает независимо от состояния утомления, так же независимо он и снижается. Таким образом, очевидно, что настройка вегетативных центров, регулирующих кровоток через мышцы, происходит в связи с утомлением во время произвольного мышечного сокращения.'
Предполагается, что эта настройка начинается путем изменения механизма обратной связи от утомленных мышц и приводит к изменению реактивности вегетативных центров, регулирующих кровоток через мышцы во время произвольных движений. Каковы бы ни были детали действующего в этом случае механизма, очевидно
ГЛАВА V
одно: что вегетативная нервная система человека претерпевает сдвиги реактивности, аналогичные явлению настройки, описанному в опытах на животных.
Барорецепторы и рефлекторная настройка гипоталамуса
В связи с важным значением гипоталамуса для организации эмоций была сделана попытка изменить его реактивность, т. е. осуществить настройку. В качестве показательной 'переменной 'было избрало кровяное давление, поскольку его изменение относительно легко вызвать в эксперименте и поскольку соответствующие опыты можно проводить иа людях.
Рецепторы, участвующие в рефлекторных изменениях возбудимости гипоталамуса при сдвигах кровяного давления, лучше всего известны по их влиянию на само кровяное давление. Это вполне логическое заключение, хотя иа первый взгляд оно кажется парадоксальным, и это следует понять прежде всего. Следующий раздел пояснит, 'каким образом сдвиг кровяного давления приводит к изменениям реактивности барорецепторов, вызывая тем самым рефлекторные компенсаторные изменения кровяного давления.
Рефлекторная регуляция кровяного давления барорецепторами
Артериальное давление представляет собою энергию (получаемую от сокращения желудочков), достаточную для проталкивания.крови через колоссальную сеть кровеносных сосудов тела. В состав этой сети входят и капилляры, сечение которых так мало, что через них с трудом проходят форменные элементы крови в один ряд. Именно через стенку капилляров осуществляется обмен веществ между кровью и тканевой жидкостью. Капиллярная кровь постоянно отдает питательные вещества и принимает продукты клеточного метаболизма. Понятно, что поддержание тока крови через капилляры имеет особое значение; застой 'быстро сделает капилляр-
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 151
ную кровь непригодной для ее роли в поддержании жидкой среды, необходимой для жизнедеятельности клеток организма.
Поскольку ткани полностью зависят от кровотока, не приходится удивляться тому, что организм выработал ряд предохранительных механизмов против его нарушений. Среди них первостепенное значение принадлежит тем, которые обеспечивают поддержание давления на соответствующем уровне. Высокая частота расстройств кровообращения, связанных с гипертонией, убедительно свидетельствует о том, что чрезмерное иовышние артериального давления не менее опасно, чем его резкое падение. Само по себе существование нормального диапазона /кровяного давления свидетельствует об эффективности.регулирующей его системы.
В норме эта система функционирует автоматически / и в значительной степени является саморегулирующейся. Как повышение, так и падение кровяного давления являются самоограничивающимися в том смысле, что и то и другое в норме довольно быстро компенсируется рефлекторными процессами, возникающими под влиянием самих сдвигов кровяного давления. Эти1 компенсаторные влияния зависят от изменения равновесия между прес-сорным и депрессорным рефлекторными механизмами. Направление этого изменения зависит от направления исходного сдвига: оно направлено так, чтобы компенсировать исходный сдвиг. Подобное компенсирующее влияние свидетельствует о наличии механизма, способного улавливать изменения,артериального давления, различать его направление и вызывать соответствующие сдвиги лрессорно-депресеориого равновесия. Таким механизмом являются барорецепторы, расположенные в стенках дуги аорты и жаротидного синуса. Эти рецепторы генерируют нервные импульсы пропорционально растяжению стенок сосудов, которое меняется в зависимости от давления находящейся в них крови. Таким образом, само кровяное давление управляет рефлекторной регуляцией нрессорно-депрессорного равновесия через барорецепторы [458].
В исследованиях, обсуждаемых в данной книге, ба-рорецепторам принадлежит ключевая роль. Необходимо
ГЛАВА V
поэтому.подробно рассмотреть механизм их действия, во-первых, в его прямой связи с кровяным давлением, во-вторых, в его зависимости от гипоталамуса и вегетативного равновесия, с которыми, по нашему мнению, неразрывно связаны эмоции и их расстройства. Прежде чем перейти к описанию действия 'барорецепторов, следует указать, что лучше других изучены 'барорецепторы, расположенные в стенках дуги аорты и каротидного синуса, которые,.пожалуй, имеют и 'наиболее важное функциональное значение, хотя и не являются единственными. Рецепторы, находящиеся в аорте, играют роль «индикаторов давления», расположенных у входа в большой «руг кровообращения. Каротидный синус — это небольшое расширение на месте отхождения внутренних сонных артерий, питающих мозг. Следовательно, 'барорецепторы занимают стратегическое положение. Поскольку барорецепторы аорты и каротидного синуса действуют одинаково, аортально-каротидные 'барорецепторы можно рассматривать как единую функциональную группу. Можно говорить также о «барорецепторных нервах», не разделяя их на каротидный и аортальный нервы. (В'близи от 'барорецепторов каротидного синуса и дуги аорты расположены хеморецопторы; подобно *барорецеп-торам, они играют важную роль в поддержании гомео-стаза, т. е. в стабилизации состояния жидких сред тела. Однако эти рецепторы не имеют отношения к обсуждаемой проблеме, и мы их здесь рассматривать не будем.)
Влияние кровяного давления на активность барорецепторов
В норме при растяжении стенок сосудов, содержащих 'барорецепторы, т. е. при каждом пульсовом толчке, по афферентным нервам барорецепторов направляются умеренные разряды (фиг. 18). Эти разряды оказывают постоянное подавляющее влияние на кровяное давление. Если кровяное давление повышается, сосуды оказываются растянутыми даже в период диастолы; в этом случае разряды являются непрерывными, но колеблются параллельно пульсовым колебаниям артериального давления.
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 153
Бронк и Стелла [109] показали, что частота разрядов одиночного «аротидного барорецептора (находится в прямой зависимости от давления в синусе. В определенном диапазоне изменения давления эти величины строго пропорциональны. Кроме того, Бронк и Стелла установили, что величина давления определяется числом возбужденных барорецепторов. Таким образом, с помощью частоты__разрядов и явления вовлечения барорецепторы точно информируют центральную нервную систему об изменениях уровня артериального давления. Наконец, эти исследователи показали, ч то адапт адия бароренеп-торов происходит очень.медленно, и поэтому они способны поддерживать относительно постоянную частоту разрядов при постоянном давлении в синусе. Таким образом, они специально приспособлены для непрерывной информации относительно уровня артериального давления независимо от того, колеблется ли оно или поддерживается на постоянном уровне.
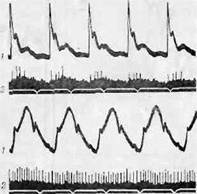
Фиг. 18. Разряд барорецепторов при низком и высоком кровяном давлении.
/ — при среднем давлении 55 мм рт. ст.] II — при среднем давлении
135 мм рт. ст.
/ — кривая артериального пульса; 2 —токи действия одиночного нервного
волокна каротидного синуса.
Влияние барорецепторов на артериальное давление
При артериальном давлении ниже 40—50 мм рт. ст. барорецепторы обычно неактивны [108]. Простой опыт, иллюстрирующий роль рефлексов с барорецепторов >в
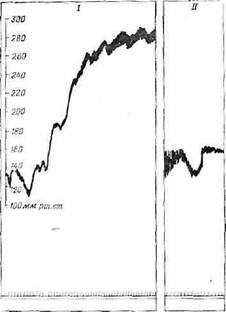
Фиг. 19. Повышение кровяного давления при перерезке барорецеп-торных нервов у собаки [456а].
/ — перерезка четырех барорецепторных нервов; II — перерезка тех же
нервов у собаки, подвергшейся полной десимпатизации. Кратковременное
начальное падение давления вызвано движением.
КТИВИОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 155
поддержании нормального кровяного давления, сводится к пережатию общих сонных артерий и регистрации сдвигов кровяного давления. При этом давление в каротидном синусе падает ниже порога возбуждения барорецепторов, а прекращение их активности приводит к быстрому, довольно значительному повышению артериального давления в большом круге. Этот эффект особенно сильно выражен, если перед пережатием •сонных артерий перерезать волокна, идущие от барорецепторов аорты. Поскольку у многих видов животных эти деп>р,ессорньге волокна идут в составе блуждающих нервов, то перерезают именно эти нервы.
Если вместо временного выключения барорецепторов пережатием сонных артерий перерезать нервы, идущие от барорецепторов афты и каротидиого синуса, кровяное давление резко повышается (фиг. 19) и остается высоким в течение месяцев и Даже лет [458].
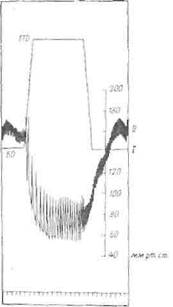
Фиг. 20. Рефлекторное падение кровяного давления и замедление сердечного ритма в результате повышения давления в изолированном каротидном синусе с сохраненной иннервацией.
/ — давление в каротидном синусе; // — частота сердечных сокращений и давление в бедренной артерии.
ГЛАВА V
Несомненно, что гипертензивные факторы обычно испытывают противодействие со стороны рефлексов барорецепторов, постоянно оказывающих депрессорное влияние, и следует иметь в виду, что ^.активност ь баро -рецепторов способствует только снижению артериального давления.
В норме повышение артериального давления усиливает разряды барорецепторов. В эксперименте не удается, изменив кровяное давление в большом -круге кровообращения, одновременно измерить влияние на него барорецепторов, активность которых изменена. Можно, однако, изолировать каротидный синус и производить его лерфузию лри любом давлении. Если каро-тидный нерв остается интактным, то влияние колебаний давления -внутри синуса л а давление в большом круге можно легко измерить. При повышении давления в синусах артериальное давление резко падает, а сердечный ритм замедляется. Аналогичный эффект дает раздражение барорецепторного нерва электрическим током (фиг. 20).
Эти эксперименты показывают, что у интактного животного повышение кровяного давления сопровождается повышением активности барорецепторов, что в свою очередь лриводит >к понижению давления.
|
|
|
|
|
Дата добавления: 2014-11-29; Просмотров: 599; Нарушение авторских прав?; Мы поможем в написании вашей работы!