КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
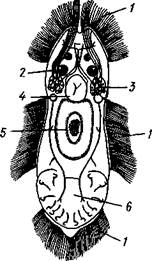
Присоски, 2 — присасывательные ямки, 3 — крючья, 4 — хоботки, вооруженные крючьями. 5 — влагалища, в которые втягиваются хоботки
|
|
|
|
Глаза, 2 — глотка, 3 — кишечник, 4 — копулятивный орг.ш, 5 — матка с яйцом, 6 — влагалище, 7 — жел-точник, 8 — яичник, 9 — семенник, 10 — прикрепительный диск с крупными срединными и мелкими краевыми крючьями, 11 — железы, 12 — зародыши 4 поколений
Рот, 2 — глотка, 3 — половое отверстие, 4 — влагалище, 5 — семяпровод, 6—матка, 7— желточный проток, S — желточники, 9 — кишечник, 10 — присоски прикрепительного диска, // — крючки прикрепительного диска. 12 — яйцевод, 13 — яичник
Рис. 149. Паразиты жабр карповых
рыб. — А — Dactylogyrus uastator (no
Быховскому и Гусеву); Б — Gyrodactylus
elegans (по Фурману):
|
| Рис. 150. Схема организации личинки (по Быховскому): / — пояса ресничек, 2 — глаза, 3 — железы, 4 — глотка, 5 — кишечник, 6 — церкомер |
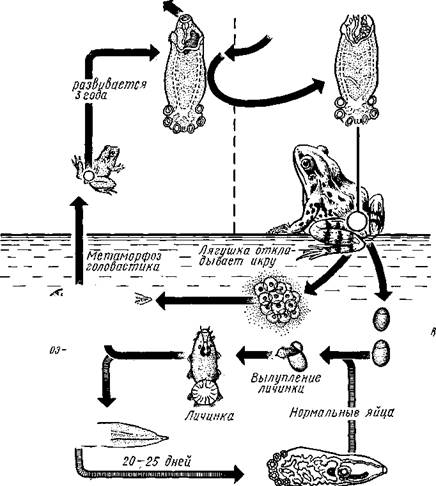
оплодотворенные яйца. Из яйца выходит маленькая мерцательная личинка, еще не имеющая на заднем конце характерных для взрослой формы шести присосок, но снабженная шестнадцатью мелкими крючьями. Личинка свободно плавает в воде, пока не прикрепится к жабрам головастика; если хозяин не будет найден, то личинка погибает. После прикрепления ресничный покров личинки сбрасывается, на заднем конце тела развиваются два крупных крючка и начинается формирование присосок. Личинки превращаются во взрослых червей и начинают откладывать яйца. Из этих яиц выходит второе поколение личинок, уже не успевающих завершить развитие на жабрах до метаморфоза хозяина, во время которого жабры атрофируются и зарастают. Личинки полистомы при этом не погибают, а по поверхности тела лягушки мигрируют в клоаку. Отсюда они забираются в мочевой пузырь, где только через три года (одновременно с лягушкой) достигают половой зрелости. В развитии Polystoma замечательна тесная связь явлений жизни паразита с отдельными периодами жизни хозяина, а также то обстоятельство, что в течение своего жизненного цикла Polystoma от эктопаразитического образа жизни (на жабрах) переходит к эндопаразитизму.
Наконец, следует остановиться на особенностях размножения еще одного представителя моногеней — Gyrodactylus (см. рис. 149,£). Представители рода интересны тем, что являются живородящими. В матке животного партеногенетическим путем развивается единственный зародыш, в котором еще до его рождения формируется зародыш следующего «внучатого поколения»; в последнем, в свою очередь, закладывается еще более молодой зародыш четвертого поколения.
Как уже указывалось, моногеней гермафродиты. В связи с этим им свойственно не только перекрестное, но и самооплодотворение. Вместе с тем есть виды, у которых при наличии гермафродитного полового аппарата выработались специальные приспособления, исключающие возможность самооплодотворения. Такими адаптациями обладает спайник парадоксальный (Diplozoon paradoxum; рис. 151), живущий на жабрах пресноводных карповых рыб. Паразит в молодости живет одиночно, но затем черви сходятся попарно и соединяются таким образом, что особая брюшная присосочка одной особи обхватывает спинной бугорок другой, и обратно. В этих местах оба животных срастаются и в таком состоянии остаются на всю жизнь. Молодой Diplozoon только в том случае и развивается дальше, если встретится с другой особью. Соединяются они таким образом, что мужские половые протоки одного экземпляра открываются в женский проток другого, чем обеспечивается перекрестное оплодотворение (рис. 151). ——————————————————————————————————————————————^
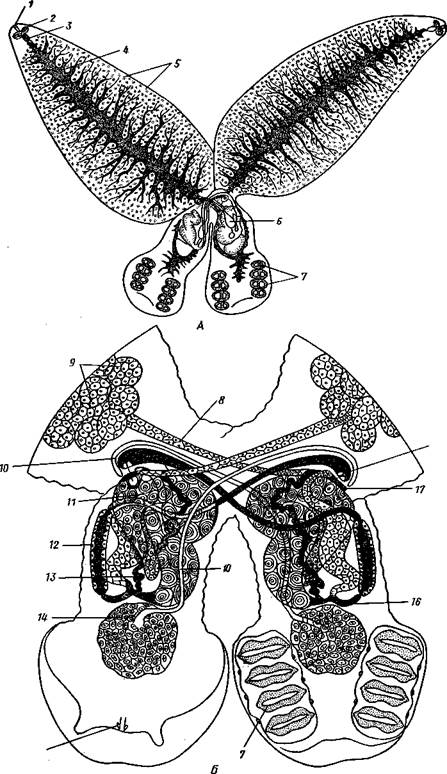
Рис. 151. Спайник парадоксальный Diplozoon paiadoxum. A — внешний вид (по Стрел-кову); Б — строение половой системы (по Целлеру):
/ — рот, 2 — ротовые присоски, 3 — глотка, 4 — кишечник, 5 — желточники, 6 — комплекс половых Желез, 7 — прикрепительные клапаны, S — желточный проток, 9 — желточники, 10 — семяпровод, 11 — яичник, 12 — матка, /3 — влагалище, 14 — семенник, 15 — крючки прикрепительного аппарата, К — яйцевод, П — место соединения семяпровода левой особи с влагалищем правой, 18 — наружное
маточное отверстие
|
| 18 |
| 15 |
Патогенное значение моногеней. Моногенеи, паразитирующие на рыбах, нередко становятся причиной их заболеваний и даже гибели, особенно в условиях прудовых рыбных хозяйств. Наиболее важное практическое значение имеют многие представители сем. Dactylogyridae, живущие на жабрах пресноводных рыб. Так, Dactylogyrus, например, нападает на молодь карпов в таком числе, что на одной рыбешке встречается до 500 этих мелких (1—3 мм) паразитов. Они питаются слизью, эпителием или (реже) кровью хозяина, вызывая иногда массовую гибель рыб. К числу опасных паразитов относятся и представители семейства Gyrodactylidae.
Гибель рыб в результате заражения моногенеями иногда происходит в природных условиях. Так, вид Nitzchia sturionis, паразитирующий на
Хозяин не размножается
Хозяин размножается
|
| I I Половые гормоны хозяина стимулируют созревание |
| Неполовозрелые полистомы |
| Личинка |
| Пол истома достигает л олово-зрелости, когда хозяин попадает 6 о од у |
| Дичинка переходит д моче -бой пузырь |
| jv^^s^iiK?4- ПрИкр'еТГ "^JtesasS ля юте я на -^ нппмп/и.нпр внутренние • ЗЖ/Р жМры fo/iee - поОеОение старших " растоо |
| Яйца выделяются. воду |
| Личинка прикрепляется к наружным кадрам молодо- €*£?^&?g. •го голоВас- •••••••••"**•• тика |
„ Жаберная форма " полистомы откладыоает яйца
Рис. 152. Ход жизненного цикла лягушачьей многоустки Polystoma integerrimum
(ВО
жабрах осетровых рыб Волги и случайно завезенный при акклиматизации севрюги в Арал, перешел там на местных осетровых (шип) и вызвал среди них массовую гибель.
Классификация моногеней основывается на особенностях строения их прикрепительного аппарата. В настоящее время Monogenoidea разбиваются но этому признаку на два подкласса, насчитывающие в общей сложности 9 отрядов.
ПОДКЛАСС I. НИЗШИЕ МОНОГЕНЕИ (POLYONCHOINEA)
К подклассу относятся моногеней, прикрепительный диск которых вооружен сильно развитыми крючьями, несет присоски или же может целиком превращаться в одну мощную присоску. Личинки (см. рис. 150) с глазами, несут на прикрепительном диске 14—16 мелких краевых крючочков. В основном—паразиты морских и пресноводных рыб. Это названные выше семейства Dactylogyridae, Gyrodactylidae и др. Представители семейства Polystomatidae паразитируют на амфибиях и рептилиях. Многие виды, например лягушачья многоустка (см. рис. 148), перешли к паразитированию во внутренних органах хозяина, т. е. стали эндопаразитами (с. 17S). Как правило, это крупные формы, характеризующиеся наличием разветвленного кишечника и множественных семенников (с. 177). Прикрепительный диск их обычно несет 6 мускулистых присосок.
ПОДКЛАСС II. ВЫСШИЕ МОНОГЕНЕИ (OLYGONCHOINEA)
Характерной чертой моногеней этого подкласса, паразитирующих только на рыбах, является наличие специализированных органов прикрепления — клапанов, действующих по принципу капкана (см. рис. 151). С появлением клапанов крючья утрачивают значение главных органов прикрепления и у взрослых червей могут иногда отсутствовать. У личинок обычно имеется 10 краевых крючочков. К этому подклассу относится рассмотренный выше Diplozoon paradoxum (см. рис. 151).
КЛАСС IV, ЛЕНТОЧНЫЕ ЧЕРВИ (CESTODA)
Ленточные черви — плоские черви, на которых паразитический образ жизни сказался значительно сильнее, чем на сосальщиках и моноге-неях. В половозрелом состоянии цестоды встречаются в кишечнике позвоночных животных; молодые стадии цестод живут в полости тела и внутри различных органов как беспозвоночных, так и позвоночных. Число видов превышает 3000.
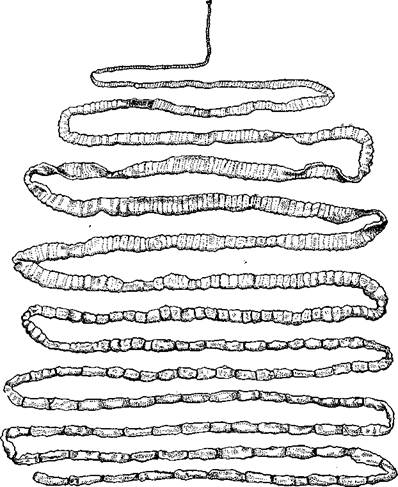
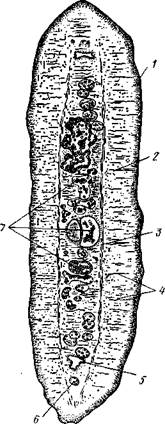
Строение и физиология взрослых ленточных червей. Тело обыкновенно бывает сильно вытянутым в длину, лентовидным и в большинстве случаев поделено па значительное число члеников, или проглоттид (рис. 153). Редко тело бывает цельным, нерасчлененным. Передний конец образует небольшую головку, или сколекс, за которым следует нерасчлененная шейка, а за ней идут проглоттиды.
Головка несет органы прикрепления, построенные по типу присосок или по типу крючков (рис. 154). Присоски имеются всегда, тогда как крючья являются менее постоянным их дополнением. Присоски устроены, в общем, так же, как у сосальщиков, и чаще всего имеются в числе четырех по краю переднего конца головки. Реже вместо типичных присосок головка снабжена двумя вытянутыми в продольном направлении щелевидными присасывательными ямками. Крючья помещаются или прямо на поверхности головки, или на особом переднем выпячивании ее — хоботке, образуя один или несколько венчиков. Хоботок втяжной. В редких случаях (Tetrarhynchus) имеется 4 хоботка, длинных, усаженных многочисленными крючками и способных глубоко вворачиваться в особые влагалища.
Проглоттиды имеют обычно четырехугольную форму, причем число их варьирует от 3 шт. до нескольких тысяч. Передние членики — самые
маленькие, по направлению же кзади размеры их постепенно возрастают. Во все время жизни червя происходит рост и увеличение числа члеников. Рост идет в области шейки: она удлиняется, и от заднего конца ее отшнуровываются все новые членики. Таким образом, самые молодые членики занимают переднюю часть тела; чем далее кзади расположен членик, тем он старее. Все тело называют цепочкой или стробилой (последнее за сходство с процессом стробиляции сцифистом сцифоидных медуз).
Величина взрослых ленточных червей колеблется между 1 мм и 10м. Окраска тела однообразная — белая или желтоватая, характерная для многих внутренностных паразитов.
| Рис. 153. Общий вид стробилы бычьего солитера (по Холодковскому) |
Цестоды обладают типичным кожно-мускульным мешком. Их покровы очень сходны с таковыми у трематод и моногеней и построены по тому же типу, что и погруженный эпителий турбеллярий. Тегумент це-стод слагается из безъядерного цитоплазматического слоя, при помощи тонких тяжей соединенного с погруженными участками цитоплазмы, несущими ядра. Отличительная особенность покровов цестод состоит в
том, что на поверхности наружного цитоплазматического слоя имеется бесчисленное множество волосковидных выростов (микротрихий), по-видимому, играющих роль в процессе питания (рис. 155). Рассмотреть их можно только с помощью электронного микроскопа. Их ультраструкту-
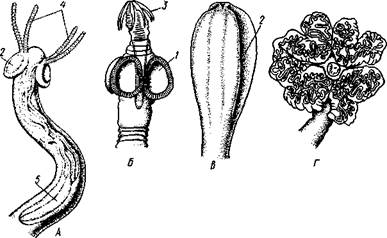
Рис. 154. Типы строения сколексов цестод (из разных авторов). А — Tetrarhynchus (Trypanorhyncha); Б — Hymenolepis (Cyclophyllidea); В — Diphyllobothrium (Pseudophyllidea); Г — Phyllobothrium (Tet-
raphyllidea):
ра принципиально отличается от таковой микроворсинок, характерных для спороцист трематод.
Непосредственно под базальной мембраной располагается наружный слой кольцевых и внутренний — продольных мускульных волокон. Часто к этим двум слоям присоединяется еще третий более глубокий слой кольцевых мышечных волокон (рис. 156). Кроме того, имеется система спинно-брюшных мышечных пучков, пронизывающих паренхиму. В последней у большинства цестод разбросаны микроскопические округлые конкреции углекислой извести — так называемые известковые тельца. Их происхождение и функция до конца не выяснены, но имеется предположение, что известковые тельца возникают как экскреты и выполняют роль своеобразной буферной системы, предохраняющей ленточных червей от вредного влияния кислой среды (например, при миграции личиночных стадий через желудок хозяина).
В паренхиме ленточных червей откладывается значительное количество гликогена, в результате анаэробного расщепления которого це-стоды (подобно трематодам) получают энергию, необходимую для жизнедеятельности.
Наиболее характерный признак ленточных червей заключается в полном отсутствии у них пищеварительной системы — факт, объясняющийся, вероятно, их образом жизни. Для цестод, обитающих в просвете кишечника другого животного (хозяина), нет необходимости самостоятельно захватывать и переваривать пищу. Последняя переводится в растворенное и доступное для усвоения состояние пищеварительными ферментами самого хозяина. Эту уже в значительной мере переваренную пищу паразиты воспринимают всей поверхностью тела. По-видимому,
особенности строения цитоплазматической пластинки тегумента, несущей микротрихии, способствуют осуществлению этого процесса. Важно отметить, что характер диеты хозяина, в особенности содержание в ней углеводов, влияет на состояние, интенсивность роста и развитие ленточных червей.
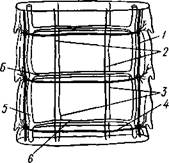
Нервная система, как и вообще у паразитов, у цестод развита слабо. Органы чувств представлены разбросанными по поверхности тела чувствительными клетками, наибольшее количество их концентрируется на сколексе. Центральная нервная система состоит из парного мозгового узла, лежащего в головке и посылающего от себя назад несколько пар нервных стволов, соединенных поперечными перемычками (ортогон). Два ствола, расположенных по бокам тела, развиты сильнее остальных (рис. 157). От стволов отходят тонкие веточки, образующие под кожей довольно густое нервное сплетение.
|
Выделительная система — протоне-фридиального типа (рис. 157, 158). По бокам вдоль всего тела, непосредственно кнутри от нервных стволов идут два главных выделительных канала. Они начинаются на заднем конце тела, затем на-
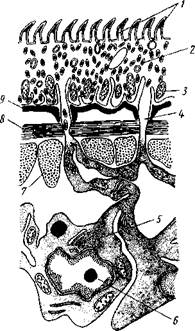
Рис. 155. Схема строения покровов цестод по данным электронной микроскопии
(по Бегину):
/ — волосовидные выросты, 2 — наружный слой цитоплазмы с митохондриями (3) и различными включениями, 4 — цитоплазматиче-ские тяжи, соединяющие наружный слой цитоплазмы с погруженной частью тегумспта, 5— погруженные клеточные тела тегумента с ядрами (6), 7—продольные мышцы, 8 — кольцевые мышцы, 9 — базальная мембрана (поперечный срез)
Рис. 156. Поперечный разрез через членик бычьего солитера Taeniarhynchus sagi-natus (по Стрелкову):
/ — кожно-мускульный мешок, 2— паренхима, 3 — слой внутренних кольцевых мышц, 4 — дорзовентральные мышцы, 5 — боковой сосуд выделительной системы, в — боковой продольный нервный ствол, 7 — перерезанные части половой системы
правляются кпереди, достигают головки, заворачиваются назад и вновь доходят до заднего конца, заканчиваясь общим выделительным отверстием. Вследствие этого часто получается впечатление, что у цестод имеются 4 продольных канала, тогда как их всего 2, но петлеобразно изогнутых у переднего конца тела. При соединении на заднем конце тела оба канала нередко образуют небольшой общий сократимый мочевой пузырь. У членистых цестод боковые каналы как в головке, так и у заднего края члеников соединяются при помощи поперечных перемычек; выделительная система получает вид лестницы. Когда последний членик цепи отвалится (вследствие периодического отрывания задних кусков стробилы), нового мочевого пузыря уже не образуется, и каждый боковой канал открывается теперь наружу особым отверстием. Многочисленные веточки каналов пронизывают паренхиму и на концах своих замыкаются звездчатыми клетками с мерцательным пламенем.
Половая система (рис. 159, 160) ленточных червей гермафродитна и, в общем, напоминает таковую сосальщиков. Лишь у некоторых из нерасчлененных цестод (Caryophylla-eus) половой аппарат одиночен. У других, например у ремнецов, имеется продольный ряд половых аппаратов, тогда как у членистых цестод в каждой проглоттиде развивается своя половая система.
Отдельные части полового аппарата у разных ленточных червей довольно сильно варьируют, так что мы для конкретности возьмем
|
Рис. 157. Часть стробилы солитера с нервными стволами и выделительными каналами (по Фурману):
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 1208; Нарушение авторских прав?; Мы поможем в написании вашей работы!