КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Кутикула 3 страница. У всех прочих брюхоногих (высшие представители подкл
|
|
|
|
У всех прочих брюхоногих (высшие представители подкл. Prosobranchia, подкл. Opisthobranchia и Pulmonata) сохраняется только одно левое предсердие, тогда как правое полностью исчезает. Положение оставшегося предсердия зависит от положения жабры или легкого. У переднежаберных и легочных оно остается впереди желудочка, а у заднежа-берных ложится позади него, так как жабра их смещается далеко назад.
Сердце обычно помещается над задней кишкой. Кроме того, оно всегда окружено перикардием, представляющим участок вторичной полости тела (целом).
|
| 1 |
| 1 |
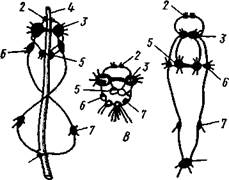
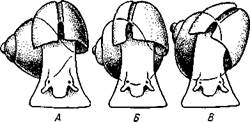
| Рис. 448. Разные формы нервной системы у брюхоногих моллюсков. А — Рго-sobranchia; Б — Opisthobranchia; В — Pulmonata (по Коршельту и Гейдеру): I — висцеральный ганглий, 1 — буккальный ганглий, 3 — церебральный ганглий, 4 — кишечный канал, 5 — педальный ганглий, 6 — плевральный ганглий, 7 — париетальный ганглий |
Периферическая часть кровеносной системы в главных чертах дает следующую картину. От желудочка отходит аорта, которая вскоре делится на два ствола: головную аорту (к переднему концу тела) и внутренностную аорту (к кишечнику, печени и половой железе). Хорошо развитая артериальная система образована сосудами с обособленными стенками. У легочных моллюсков более крупные сосуды постепенно переходят в капилляры. В конце концов, однако, кровь изливается в мелкие лакуны соединительной ткани, теряет кислород и постепенно собирается в более крупные венозные лакуны, наиболее развитая из них омывает желудок, печень и гонаду. Отсюда кровь возвращается к органам дыхания (жабрам или легкому) и, окислившись, попадает непосредственно в сердце. Таким образом, сердце брюхоногих наполняется артериальной кровью.
Кровь чаще всего бесцветна и содержит амебоциты. Иногда в крови имеется вещество, близкое к гемоглобину. У некоторых брюхоногих удалось обнаружить присутствие в крови белковых соединений, содержащих марганец, который у них играет, по-видимому, такую же роль, как железо в гемоглобине крови позвоночных.
Нервная система брюхоногих (рис. 448) хорошо развита и представляет сравнительно-анатомический интерес. В общем, от низших брюхоногих к высшим наблюдается постепенный переход от системы, напоминающей таковую подтипа Amphineura, к разбросанно-узловой нервной системе из нескольких ганглиев, соединенных между собой свободными от нервных клеток волокнистыми перемычками. У наиболее примитивных брюхоногих моллюсков ганглии либо отсутствуют, либо выражены еще очень слабо. Нервные клетки в этом случае располагаются по ходу стволов. Последующие изменения происходят путем концентрации ганг-лиозных клеток в некоторых точках нервных стволов, в которых и образуются ганглии. У переднежаберных моллюсков первично обособляется пять пар основных ганглиев.
1. Над глоткой расположены два церебральных ганглия, соединенных церебральной комиссурой.
2. В ноге педальные стволы концентрируются в передней части в два педальных ганглия, соединенных под глоткой педальной комиссурой. Кроме того, они связаны с церебральными ганглиями двумя продольными перемычками, или коннективами (напомним, что поперечные нервные перемычки, соединяющие одноименные ганглии, принято называть ко-миссурами, продольные перемычки между ганглиями разного наименования— коннективами). У ряда примитивных современных форм обра-
зования педальных ганглиев еще не произошло, и вместо них сохраняются 2 педальных ствола.
Далее на пути плевровисцеральных стволов обособились еще три пары ганглиев.
3. Два плевральных ганглия расположены приблизительно на уровне педальных ганглиев; они соединены коннективами с церебральными и педальными узлами.
4. Далее кзади на пути плевровисцеральных стволов залегают два париетальных ганглия.
5. Наконец, еще дальше, под задней кишкой, лежит пара висцеральных ганглиев, соединенных друг с другом висцеральной комиссурой.
Последние три пары ганглиев как бы нанизаны на ту нервную петлю, которая образуется у подтипа Amphineura плевровисцеральными стволами. Участок стволов, находящийся между плевральным и висцеральным узлами каждой стороны и разбитый париетальным ганглием на 2 коннектива, называется плевровисцеральным коннективом.
Кроме этих основных ганглиев образуются еще дополнительные ганг-лиозные массы, иннервирующие отдельные органы (например, глоточный, или буккальный ганглий; рис. 448). У заднежаберных (подкл. Opistho-branchia) и легочных (подкл. Pulfnonata) моллюсков первично обособляется не 5, а 7 пар ганглиев. У всех моллюсков эти цифры могут уменьшаться за счет слияния ганглиев в более сложные нервные узлы.
Своеобразные изменения нервной системы брюхоногих моллюсков связаны с появлением у них асимметрии. Происхождение этих изменений подробнее будет рассматриваться дальше (с. 465). Главная суть их в следующем. У всех переднежаберных и немногих низших представителей двух других подклассов плевровисцеральные коннективы образуют перекрест так, что находящиеся на их протяжении париетальные ганглии перемещаются: левый на правую сторону (под кишку), а правый — на левую сторону тела (над кишечником). Нервную систему таких брюхоногих называют хиастоневральной или перекрещенной (рис. 448). У основной массы легочных и заднежаберных моллюсков перекрест коннективов вторично исчезает, но правый плевровисцеральный коннек-тив часто заметно короче левого. Это брюхоногие с неперекрещенной, или эутиневральной, системой. У высших Gastropoda плевровисцеральные коннективы вообще настолько укорачиваются, что ганглии собираются в одну общую массу вокруг глотки.
Церебральными ганглиями иннервируются глаза, статоцисты, глотка и головные щупальца, педальными — мускулатура ноги. Плевральные ганглии снабжают нервами главным образом мантию. Париетальными ганглиями иннервируются ктенидии и осфрадии; наконец, висцеральными ганглиями осуществляется иннервация внутренностей.
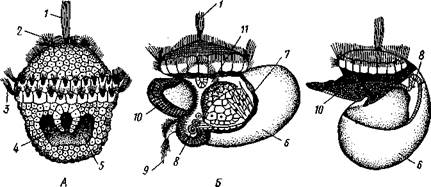
Органы чувств. Для осязания служат головные щупальца (см. рис. 438; 443), края мантии и некоторые другие места. Кроме того, имеются органы химического чувства. Ими считают прежде всего осфрадии, лежащие у основания ктенидиев. В наиболее развитом состоянии они имеют форму продолговатого валика, лежащего у основания жабры и.но обеим своим сторонам несущего 100—150 листочков. Вследствие этого осфрадии иногда очень похож по виду на жабру. Внутренность валика занята скоплением ганглиозных клеток, от которого отходят нервы к листочкам.
Передняя пара головных щупалец, называемых губными, тоже, по-видимому, служит для восприятия химических раздражений, исполняя роль органов вкуса и обоняния.
Органы равновесия в виде пары статоцистов имеются у всех Gastropoda. Это обыкновенно два замкнутых пузырька, эпителий которых состоит из мерцательных и чувствительных клеток (рис. 449); последние иногда образуют на стенке пузырька особое скопление — «слуховое пятно». В жидкости, заполняющей пузырек, плавают от 1 до 100 конкре-
|
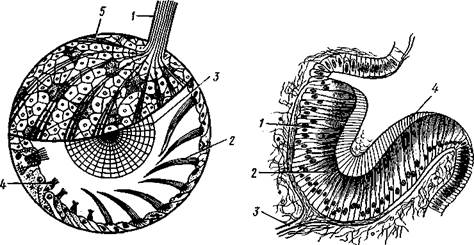
Рис. 449. Статоцист киленогого моллюска Pterotrachea (по Бючли): / — нерв. 2 —пучок ресничек, 3 — статолит, 4 — чувствительные клетки, 5 — нервные волокна
Рис. 450. Разрез глаза морского блюдечка Patella rota (по Гильгер):
J — сетчатка, 2 —пигментированная зона, 3 —• зрительный нерв, 4 — утолщенная кутикула
ций углекислой извести — статолитов, которые играют роль слуховых камешков. Различное положение их в статоцистах и надавливание то на одни, то на другие чувствительные клетки позволяют животному ориентироваться в пространстве. Оба статоциста лежат всегда в тесном
|
соседстве с педальными ганглиями, но иннервируются они тем не менее от церебральных ганглиев при помощи двух нервов.
Одна пара глаз имеется почти у всех брюхоногих; они лежат на голове у основания, а иногда на вершине задней пары щупалец, соответственно часто обозначаемых как глазные. Сложность строения глаз варьирует от поо-стых ямок до глазных пузырей с хрусталиком и стекловидным телом (рис. 450, 451).
| чаще сохраняется лишь одна левая (рис. 452). Одним концом почки посредством ресничной воронки сообщаются с перикардием, |
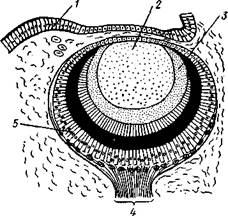
Выделительная система брюхоногих состоит из пары почек типа целомодуктов, из которых
Рис. 451. Разрез глаза Fissurclla (из Догеля):
/ — кожный эпителий головы, 2 — хрусталик,
3 —стекловидное тело, 4 —зрительный нерв,
5 — зрительные клетки (сетчатка)
|
т. е. с участком целома, другим — открываются в мантийную полость сбоку от порошицы. Две почки имеются только у низших представителей подкл. Prosobranchia, но одна из них развита слабее другой.
| Рис. 452. Выделительные органы брюхоногих моллюсков. А — мочеполовая система Puncturella noachina (Prosobranchia Dioto-cardia) (из Иванова); Б— почка и сердце улитки Daudebardia rufa (Pulmonata) (no Лангу): 1 и 9 — наружные почечные отверстия, 2 — левая почка, 3 — правая почка, 4 — половая железа, 5 — проток половой железы, 6 — канал почки, открывающийся в перикардий. 7 — отверстие ппчки в перикардий, 8 — мочевая камера, 10— перикардий с сердцем, // — мочеточник, 12 — задняя кишка |
Половая система обнаруживает у брюхоногих большие вариации (переднежа-берные обычно раздельнополы, легочные и заднежа-берные — гермафродиты). Низшие брюхоногие не имеют специальных половых протоков, и половая железа открывается у них в правую почку.
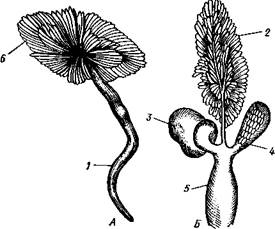
|
Половая железа всегда одна. У раздельнополых форм (рис. 453) это яичник или семенник, у гермафродитов — гермафродитная железа, в которой образуются и живчики и яйца. Выводные протоки Prosobranchia сравнительно просты. Самец имеет семяпровод, открывающийся наружу близ переднего конца тела на правой стороне. Либо у самого отверстия, либо впереди него на голове помещается мускулистый вырост — совокупительный орган.
У самки яйцевод может образовывать местное расширение — матку, а также семяприемник.
| Рис. 453. Половой аппарат самца (А) и самки (Б) киленогого моллюска Pterotrachea (из Клауса): / — семяпровод, 2 — яичник, 3 — белковая железа, 4 — семяприемник, 5 — матка, 6 — семенник |
Половые пути Pulmonata и Opisthobranchia несравненно сложнее, как можно видеть на примере виноградной улитки (см. рис. 446). От гермафродитной железы отходит общий гер-мафродитный проток, принимающий в себя выводной канал особой белковой железы. После впадения в проток железы он расширяется, причем главная часть его просвета выполняет роль яйцевода, а узкий желоб, проходящий по одной стороне яйцевода, проводит семя. Далее этот общий проток разделяется на два самостоятельных канала: яйцевод и более тонкий семяпровод. Семяпровод переходит в мускулистый совокупительный
орган (пенис). Яйцевод расширяется и образует матку, в которую впадают протоки пальцевидных желез. Матка при помощи влагалища открывается в особое впячивание стенки тела — половую клоаку, куда открывается и мешок совокупительного органа. Кроме того, во влагалище впадают еще два мешковидных образования — семяприемник, предназначенный для восприятия семени другой особи, и мешок с «любовной стрелой»; стрела — это игла из углекислой извести, которая при копуля-
|
| в |
Рис. 454. Развитие морского блюдечка Patella (Prosobranchia, Diotocardia) (по Паттен). А — трохофора; Б — парусник до скручивания; В — парусник
после скручивания:
1 — теменной султан, 2 — реснички верхнего полушария, 3 — прототрох, 4 — рот, 5 — просвечивающие мезодермальные полоски, 6 — раковина, 7 — внутренностный мешок, в — мантия, 9 — реснички задней части тела, 10 — зачаток ноги, 11 — кишечник
ции втыкается в кожу другой особи и служит для ее раздражения. Оплодотворение перекрестное.
Развитие. Среди примитивных брюхоногих встречаются формы, обладающие наружным оплодотворением, у большинства же форм оплодотворение внутреннее. Яйца часто окружаются пергаментными или студенистыми коконами и соединяются в кладки. Дробление яйца чрезвычайно напоминает таковое Polychaeta (с. 261). Оно полное, неравномерное, детерминированное, протекает по спиральному типу. Имеются те же макромеры А, В, С и D с четырьмя квартетами микромеров. Главная часть мезодермы тоже формируется за счет клетки D — из ее потомка 4d. Целомическая мезодерма образует две мезодермальные полоски, происходящие от двух первичных мезодермальных клеток. У низших Prosobranchia из яйца развивается настоящая трохофора с предротовым венчиком ресниц (прототрохом) и с теменным султаном (рис. 454, Л). Затем трохофора превращается в личинку парусник, или велигер (рис. 454, В). У большинства представителей подкл. Prosobranchia и Opisthobranchia из яйца выходит сразу парусник. Парусник часто характеризуется развитием на предротовом отделе тела 2—4 боковых лопастей, окаймленных предротовым венчиком ресниц. Лопасти образуют мерцательный парус, от которого и сама личинка получила свое наименование.
Парусник обнаруживает ряд признаков, специфичных для моллюсков. На его спинной стороне появляется утолщение эктодермы, которое затем углубляется и дает легкое впячивание — раковинную железу. На дне впячивания выделяется нежный роговой зачаток раковины; впячивание затем выворачивается, зачаток раковины занимает поверхностное
|
| 8 |
положение и разрастается. На брюшной стороне тела, позади рта личинки, образуется выступ, нога, на которой в виде двух маленьких боковых впячивании эктодермы закладываются статоцисты. Впереди рта появляются щупальца и глаза. Позади зачатка раковины эктодерма образует складку, нависающую над порошицей и прикрывающую зачаток мантийной полости. У легочных мантийная складка продолжает расти вперед и становится глубже лишь на правой стороне тела, постепенно перемещаясь к переднему концу. У низших брюхоногих вместо этого происходит быстрый поворот зачатка мантийной полости с порошицей и раковины на 180° (рис. 454, Б, В), вследствие чего порошица ложится на переднем конце туловища, позади головы. Свободноплавающий парусник испытывает в конце превращения редукцию предротового венчика ресниц, опускается на дно и переходит к ползающему образу жизни. Описанный метаморфоз характерен для большинства представителей подклассов Prosobranchia и Opisthobran-chia; у Pulmonata развитие прямое и протекает до конца внутри яйцевой оболочки.
| 10 |
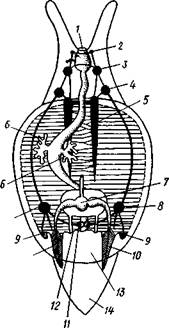
Происхождение асимметрии. Ряд ученых старались разгадать происхождение асимметрии у брюхоногих, но часть высказанных гипотез ныне имеет лишь исторический интерес.
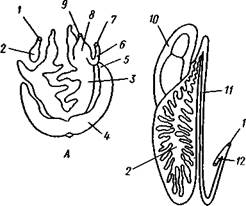
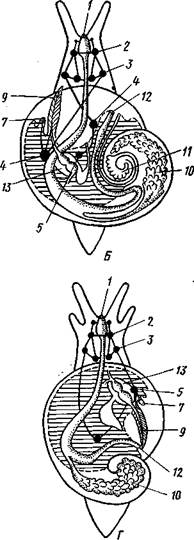
| Рис. 455. Гипотетический симметричный предок Gastropoda (по Штемпелю): 7 — рот, 3 — буккальный ганглий, 3 — церебральный ганглий, 4 — плевральные ганглии, 5 — педальный ствол, 6 — печень, 7 — желудочек сердца, 8 — париетальные ганглии, 9 — осфрадии, 10 — ктенидии, // — висцеральные ганглии, 12 — отверстие почки, 13 — мантийная полость, 14 — задний конец ноги |
Наиболее современной и обоснованной является теория Нэфа (1913). Есть все основания предполагать, что предками класса Gastropoda были вполне симметричные моллюски с порошицей и мантийным комплексом на задней стороне тела (рис.455), но имели раковину, закрученную в одной плоскости. Такая раковина нисколько не нарушала билатеральной симметрии. Так как вход в мантийную полость, а следовательно, и устье раковины находились сзади, то завиток раковины помещался впереди, над
головой. Такая раковина несвойственна современным брюхоногим, но она характерна для примитивного головоногого моллюска — кораблика (Nautilus, с. 486). Кораблик ведет плавающий образ жизни, его раковина с завитком, обращенным вперед и закрученным в одной плоскости, не препятствует плаванию. При ползании по дну такая раковина, напротив, была бы значительной помехой, завиток ее давил бы на голову, а край устья скреб бы дно. Поэтому естественно допустить, что предок брюхоногих вел плавающий образ жизни.
Причиной появления характерных черт организации Gastropoda, по Нэфу, послужил переход их предка от плавающего к ползающему образу жизни. При этом сразу же проявились отмеченные неудобства рако-кины с завитком, обращенным вперед. Устранение этих неудобств происходило следующим образом. Характерной особенностью организации брюхоногих и их ближайших предков является разделение тела на 2 области — голову с ногой и внутренностный мешок, связанные узким
|
|
|
мускулистым стебельком — основанием внутренностного мешка. Относительное положение главных частей тела может значительно изменяться благодаря мышечным сокращениям стебелька. Это прекрасно видно на современных брюхоногих, способных поворачивать раковину с лежащим внутри нее внутренностным мешком. При этом голова и тесно связанная с ней нога сохраняют свое нормальное положение, а внутренностный мешок может мышечными усилиями поворачиваться вокруг вертикальной оси на 180° и более. Такие временные повороты внутренностного мешка на стебельке называются физиологической тор-сией (скручиванием).
| Рис. 456. Схема происхождения переднего положения мантийного комплекса и хиастонсв-рии. Верхний ряд — вид сбоку, нижний — со спинной стороны (по Нэфу). А— исходная форма со спиральной симметричной раковиной, обращенной вершиной вперед; Б — промежуточная стадия; В — форма с законченным скручиванием |
Перейдя к ползанию по дну, предок Gastropoda вынужден был повертывать внутренностный мешок с раковиной (рис. 456). Поворот на 180° оказался наиболее выгодным. Сначала он носил характер временной физиологической торсии, но потом новое положение раковины — завитком назад, а устьем вперед — стало постоянным и закрепилось наследственно. Именно такое положение раковины и характерно для современных представителей кл. Gastropoda. Замечательно, что молодой парусник
низших переднежаберных имеет сначала раковину, обращенную завитком вперед (см. рис. 454,5), мантийные органы развиваются у него на задней стороне тела. Затем, опустившись на дно и переходя к ползанию, он испытывает торсионный процесс — вследствие сокращения мышц раковина с внутренностным мешком сразу (иногда в течение нескольких минут) поворачивается на 180° вершиной назад (см. рис. 454, В). У высших форм класса Prosobran-chia этот процесс происходит на более ранних стадиях развития и сводится к неравномерному росту: левая сторона основания внутренност-

|
|

|
| Б — |
Рис. 457. Раковина Tremanolus (Bellerophontidae) (из Мура). А — снизу;
сверху; В — сбоку
ного мешка растет быстрее правой. Поэтому у них из яйца выходит парусник с раковиной, обращенной вершиной назад и с передним положением мантийного комплекса.
Итак, у первичных древних брюхоногих раковина повернулась на 180°. Мантийные органы при этом оказались над головой на передней стороне туловища, а длинные плсвровисцеральные коннективы неизбеж-
|
|
Рис. 458. Схема перехода к асимметричной па-ковине. Вид спереди (по Нэфу). А и Б — последовательные стадии: / — ктенидии, 2 — желудочек сердца, 3 — предсердия. 4 — задняя кишка
Рис. 459. Схема регуляции положения раковины. Вид спереди (по Нэфу). А — первичное состояние, ось раковины поперечна; Б — ось раковины занимает косое положение, вершина сдвинута назад; В — нормальное положение раковины, вершина приподнята кверху
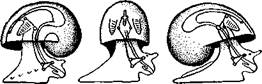
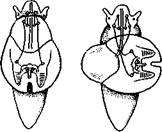
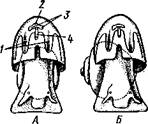
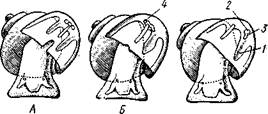
но перекрестились, т. е. возникла хиастоиеврия (рис. 456). Раковина, повернувшаяся вершиной назад, сперва оставалась вполне симметричной. Именно такими брюхоногими, по-видимому, были ископаемые Bel-lerophontidae из нижнего палеозоя (рис. 457). Следовательно, раковина, повернутая вершиной назад, переднее положение мантийного комплекса и хиастоневрия возникли одновременно как результат единого процесса филогенетической торсии.
|
| В |
Дальнейшие эволюционные изменения в пределах класса Gastropoda сводятся к следующему. В результате ползающего образа жизни увеличилась нога и соответственно должен был возрасти объем раковины, в которую при опасности втягиваются как голова, так и нога. Увеличение объема раковины при сохранении прежнего ее диаметра возможно лишь за счет расположения ее оборотов в разных плоскостях. Поэтому раковина приняла коническую форму, а вершина ее все более и более стала выдаваться вправо (рис. 458). Но такая асимметричная (правозакручен-ная) раковина имеет неустойчивое положение, перетягивая животное на правый бок. По-этому для сохранения равновесия онг все более отклоняется вершиной назад и вверх, а устье ее повертывается несколько вправо (рис. 459), г. е. происходит некоторое раскручивание, или деторсия. Это яв- „... „
Рис. 460. Схема образования асимметрии ман-
ление, называемое регуляцией ТИЙНОГо комплекса. Вид спереди (по Нэфу).
положения раковины, приво- А — В — стадии постепенной редукции правого
дит к следующему: 1. Внут- ктенидия и предсердия и смещения задней
ревностный мешок давит своей кишки направо:
пприл;М7Тттрртприил i-n / —ктенидии, 2 — желудочек сердца, 3 — левое пред-
Преимущественно ha сердне, < — рудимент правого предсердия
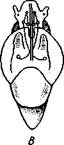
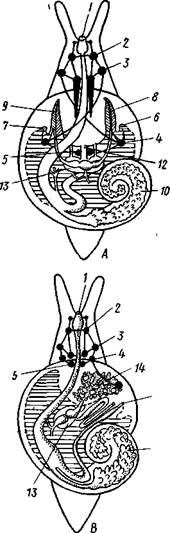
правую сторону мантийной полости. 2. Недостаток места в правой части мантийной полости и повышенное давление вызывают постепенную редукцию и, наконец, полную утрату правых мантийных органов (рис. 460). Так возникают характерные черты Prosobranchia Monoto-cardia (рис. 461), которые лишены правых ктенидиев, осфрадия, предсердия и почки.
|
|
| 12 |
| 10 |
Рис. 461. Схема типов асимметрии у разных Gastropoda (по Штемпелю). А — Prosobranchia Diotocardia; Б — Prosobranchia Monotocardia; В — Pulmonata; Г — Opisthobranchia:
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 539; Нарушение авторских прав?; Мы поможем в написании вашей работы!