КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Стебель. Почка – это зачаточный побег
|
|
|
|
Почки
Почка – это зачаточный побег.
Почки растений могут быть вегетативными, несущими только зачаточные стебли и листья, генеративными (от лат. «genero» – рождаю, произвожу), несущими только зачаточный цветок или соцветие, и смешанными.
По местоположению и функциям почки могут быть верхушечными (расположенными на верхушке оси любого порядка), боковыми (развивающимися в пазухах листьев, дающими побеги следующего порядка) и придаточными, или адвентивными (возникающими на взрослых частях органов из внутренних тканей, обеспечивающих вегетативное возобновление).
У некоторых растений придаточные почки формируются на корнях (корнеотпрысковые растения – малина, одуванчик, иван-чай), на листьях (выводковые почки у бриофиллюма).
Распространенный тип почек у древесных или травянистых многолетних растений, являющийся их обязательным признаком, – почки возобновления, впадающие на некоторое время в состояние покоя.
Почки и побеги обогащения развертываются одновременно с ростом материнской оси, обеспечивают обильное ветвление.
Спящие почки ежегодно нарастают в толще ствола древесных растений, образуя скрытую веточку, которая может ветвиться. Таким образом разрастается, например, пневая поросль. Спящие почки плохо формируются у хвойных растений.
По наличию или отсутствию почечных чешуй почки могут быть закрытыми (имеющими почечные чешуи) и открытыми (без почечных чешуй). В обоих случаях защита почек осуществляется за счет опушения, высокого осмотического давления, наличия дубильных веществ, склеивания почечных чешуй.
Чтобы занимать в почке возможно меньше места и в тоже время хорошо защищать стеблевую верхушку, молодые листья в генеративной или смешанной почке находятся в свернутом положении, причем способы этого сворачивания, называемого листосложением, бывают различными. Основные типы листосложения отражены на рис. 4: сдвоенное (а) – у шиповника; складчатое (б) – у ольхи; скомканное (в) – у сныти; свернутое (г) – у вишни; завернутое (д) – у тополя; отвернутое (е) – у клюквы.
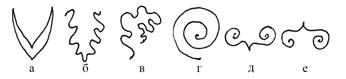
Рис. 4. Типы листосложения
Стебель растения – осевая часть побега, растущая за счет верхушечного и вставочного роста. В отличие от корня в росте стебля отсутствует строгая акропетальность (верхушечное нарастание). На апексе побега с правильной периодичностью возникают листовые зачатки, что обусловливает раннее развитие узлов. Рост междоузлий запаздывает. Междоузлия растут и развиваются позже – за счет работы интеркалярных меристем в основаниях молодых междоузлий.
Функции стебля: опорная, проводящая, запасающая, фотосинтетическая.
Главный стебель дерева – ствол. Отдельные крупные стебли у кустарников – стволики.
По форме поперечного сечения (рис. 5) стебель может быть округлым (а), сплюснутым (б), трехгранным (в), четырехгранным (г), многогранным (д), крылатым (е).
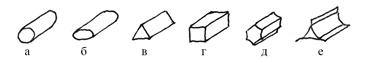
Рис. 4. Виды стеблей по поперечному сечению (см. в тексте)
По положению в пространстве стебель может быть приподнимающимся (напр., у клевера лугового), прямостоячим (у коровяка черного), ползучим (у вербейника монетного), вьющимся (у вьюна полевого), цепляющимся (у горошка мышиного).
В первичном строении стебля выделяют первичную кору и стелу. В состав первичной коры входят хлоренхима, механические ткани, неспециализированная паренхима, выделительные и другие ткани. Сверху первичная кора покрыта не входящей в ее состав эпидермой с устьицами.
Стела (от греч. «стеле» – столб) – осевой цилиндр стебля. Первичные проводящие ткани часто разделены на проводящие пучки. Между пучками находятся клетки паренхимы. Иногда разделение на пучки выражено неотчетливо (у сливы) или отсутствует (у подмаренника). Первичная ксилема находится рядом с сердцевиной, первичная флоэма – рядом с корой. Между первичной флоэмой и корой обычно развиваются механические ткани (хорошо выраженные по периферии и отсутствующие в центре).
Внутрь от проводящих тканей – сердцевина из неспециализированной паренхимы. Сердцевина может содержать запас веществ, идиобласты, воздушную полость.
Общий план строения стебля первичного строения представлен на рис. 6.
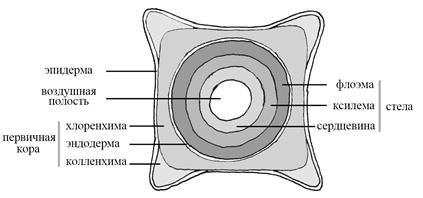
Рис. 6. Стебель подмаренника душистого
Единая проводящая система побега закладывается в апексе: через узел в стелу побега внедряются растущие прокамбиальные пучки развивающегося листа. Проводящие ткани листа продолжаются непосредственно в стебель.
Совокупность всех пучков листа, входящих в узел, называется листовым следом. Пучки проходят в стелу через паренхиму узла. В ней формируются листовые лакуны. Соответственно, листовые следы могут быть одно-, двух-, многопучковыми, а также одно-, двух- и многолакунарными.
Принципы строения и взаимоотношениях между типами стелы у высших растений легли в основу стелярной теории. Существует несколько основных стелярных типов, характерных для крупных групп высших растений, и эволюционная преемственность этих типов. При микроскопировании поперечных срезов стеблей растений тип стелы является одним из наиболее ярких диагностических признаков для отнесения исследуемого растения к крупным таксономическим группам.
На рис. 7 приведены основные типы стел и их эволюционная взаимосвязь.
Гаплостела (от греч. «гаплос» – простой) представляет сплошной тяж ксилемы, покрытый слоем флоэмы. Такой тип стелы свойственен риниофитам (вымершая группа растений), а также, напр., тычиночным нитям покрытосеменных растений.
Актиностела (от греч. «актис» – луч) происходит от гаплостелы. Ксилема на поперечном срезе имеет форму звезды, закладывается экзархно, развивается центростремительно. Флоэма отчасти повторяет форму звезды, окружая ксилему, но преимущественно развивается между ее лучами. Переход от гапло- к актиностеле обусловлен появлением пучков, идущих в боковые органы побега (листья). Такой тип стелы характерен для плауновидных. У производной актиностелы – плектостелы, также встречающейся у некоторых плаунов, флоэма не только окружает тяжи ксилемы снаружи, но и есть в промежутках между ними, что на поперечном срезе придает стеле слоистую структуру.
Сифоностела (от греч. «сифон» – трубка) происходит от гаплостелы и характеризуется появлением сердцевины, что дает возможность существования более крупных форм растений, т. к. перемещение прочной ксилемы на периферию стебля и образование трубчатой конструкции делают стебель более прочным. Паренхимная сердцевина обеспечивает запас веществ. Такой тип стелы свойственен, напр., подмареннику.
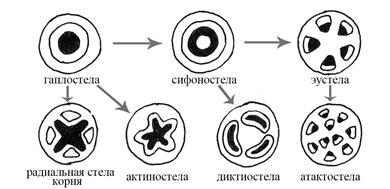
Рис. 7. Типы стел
Диктиостела (от греч. «диктион» – сеть) происходит от сифоностелы. Дальнейшее ее усложнение связано с развитием крупных листьев и боковых побегов: сифоностела рассекается паренхимой на крупные отдельные пучки. На поперечном срезе образуется прерывистое кольцо. Данный тип стелы встречается у папоротников.
Эустела (от греч. «эу» – истинный) происходит от сифоностелы. Составлена коллатеральными открытыми пучками, характерна для двудольных растений. Разновидностью эустелы является артростела (от лат. «arthrus» – членистый), встречающаяся у хвощевидных. В случае артростелы в узле каждый проводящий пучок расщепляется на три веточки, из которых одна идет в лист, а две другие соединяются в следующем узле с соседними веточками, что формирует членистую проводящую сеть.
Атактостела (от греч. «а» - отрицание; «тактос» – расположение в порядке) происходит от эустелы, характерна для однодольных растений. У этого типа стелы пучки закрытые, распределены по всему поперечному сечению стебля. Часто в стеблях однодольных бывает крупная центральная воздушная полость. Такой тип стебля однодольных называется соломиной. В этом случае проводящие пучки атактостелы смещены к периферии стебля.
Вторичное утолщениестебля в первую очередь связано с работой камбия. Он может возникать как непрерывный слой (или кольцо) в прокамбии и длительно откладывать сплошные слои вторичной ксилемы (древесины) и флоэмы (луба).
Камбий состоит из тонкостенных клеток веретеновидной формы, заостренных на концах и вытянутых вдоль оси стебля. Плоские широкие стороны обращены к флоэме (наружу) и кселеме (внутрь). Остальные клетки смыкаются с соседними клетками камбия. Совокупность похожих на клетки камбия клеток – это камбиальная зона.
Деление клеток камбия происходит тангенциально (параллельно плоским сторонам клеток). В сторону ксилемы обычно откладывается больше клеток, чем в сторону Фл. Поэтому вторичная древесина нарастает быстрее луба.
Среди основных (веретеновидных) клеток камбия возникают группы коротких инициальных клеток, дающих начало лубодревесинным лучам (вторичным сердцевидным лучам). Это – лучевые инициали, возникающие путем поперечного деления веретеновидных инициальных клеток камбия.
У древесных семейств растений камбиальное утолщение идет длительные годы. В центре ствола формируется массив древесины (до 90 % всего объема стебля). Основная ее часть составлена клетками без живого содержимого (сосуды, трахеиды, волокна). Среди них присутствуют живые элементы лучевой и вертикальной парехнимы. Образуется связанная система для запасания и передвижения запасных веществ. Древесина выполняет свою водопроводящую функцию только до тех пор, пока в ней содержатся живые элементы.
Весной камбий образует преимущественно водопроводящие элементы – сосуды и проводящие трахеиды. Летом образуются узкие элементы с толстыми стенками – волокнистые трахеиды и древесинные волокна. Переход от весенней древесины к летней – постепенный, а от летней к весенней следующего года – резкий. Образуется годичное кольцо.
У некоторых растений образуются тилы – выросты живых клеток, внедряющиеся в полость сосуда и закупоривающие его. В тилах накапливаются смолы и дубильные вещества. Это предохраняет многолетние стебли, содержащие много мертвых элементов, от гниения.
Вторичная кора растений образована вторичной флоэмой (лубом), остатками первичной флоэмы и первичной коры и перидермой. Из наружных слоев вторичной коры развивается третичная ткань – корка.
Луб также состоит из элементов двух систем – вертикальной и горизонтальной. К вертикальной системе луба относятся ситовидные элементы, клетки-спутницы, вертикальные тяжи лубяной паренхимы и лубяные волокна. К горизонтальной (радиальной) системе луба относятся лубяные лучи.
Вторичная кора по мере утолщения стебля деформируется в двух направлениях: растягивается по окружности, что вызывает разрастание клеток паренхимы первичных лучей, и сдавливается в радиальном направлении, из-за чего флоэма перестает через год проводить вещества. Функция мертвых элементов флоэмы – защита глубже расположенных участков стеблей растений.
Массовый транспорт веществ идет по молодым слоям луба и древесины (около камбия). Луб теряет проводящую способность через год, древесина – через несколько лет. Водопроводящий (живой) слой древесины называется заболонь. Остальной массив древесины, потерявший способность к водопроведению, называется ядром и выполняет опорную функцию.
К особенностям строения древесины голосеменных растений можно отнести следующие. Основная масса древесины составлена трахеидами длиной до 4 мм. Располагаются правильными радиальными рядами. В ранней древесине трахеиды тонкостенные, с хорошо выраженными порами (водопроводящие), в поздней – толстостенные с узкими полостями и порами (опорные). Граница между годичными слоями выражена очень резко.
Древесинные лучи голосеменных растений также имеют некоторые особенности, диагностические при определении древесины растений под микроскопом: клетки древесинного луча расположены в несколько ярусов, что хорошо заметно на тангенциальном срезе. Верхний и нижний ярусы состоят из мертвых клеток с окаймленными порами – это лучевые трахеиды, функция которых – водопроведение. В среднем ярусе находятся живые клетки, в которых накапливаются и передвигаются пластические вещества.
В древесине голосеменных семейства сосновых также имеется система вертикальных и горизонтальных смоляных каналов, различимых при микроскопировании по хорошо заметным мелким обкладочным клеткам, окружающим округлые в сечении смоляные канальцы.
В стволах хвойных деревьев вода движется медленнее, чем у лиственных пород (т. е. у покрытосеменных древесных растений). Это явилось одной из причин быстрого расцвета покрытосеменных и расселения их на больших пространствах.
|
|
|
|
|
Дата добавления: 2015-04-25; Просмотров: 3347; Нарушение авторских прав?; Мы поможем в написании вашей работы!