КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Гликолиз. Брожение
|
|
|
|
Переваривание углеводов
Углеводы составляют большую часть пищи человека, около 60-70% пищевого рациона. В среднем количество углеводов в суточном рационе человека составляет 450—600 г. Организм человека и животных получает углеводы с различными пищевыми веществами, главным образом, растительного происхождения. Окисление углеводов в тканях является одним из основных источников энергии, необходимой организму для осуществления разнообразных функций. Избыток углеводов в пище ведет к превращению их в жиры. Избыточное введение углеводов может привести к нежелательному ожирению у человека. Недостаточное потребление углеводов для человека также нежелательно. Оно может закончиться нарушением обмена веществ. Распад углеводов начинается в ротовой полости. В слюне содержится фермент, называемый µ-амилазой, расщепляющий крахмал. В желудке углеводы не подвергаются перевариванию, так как там нет соответствующего фермента. Основное переваривание углеводов происходит в двенадцатиперстной кишке и в дальнейших отрезках тонких кишок под влиянием µ-амилазы, поступающей в двенадцатиперстную кишку с соком поджелудочной железы. Главным, конечным продуктом гидролиза крахмала µ-амилазой является мальтоза, которая затем расщепляется на две молекулы глюкозы под действием фермента мальтазы. Мальтаза расщепляет дисахариды до моносахаридов. Сахараза гидролизует сахарозу на глюкозу и фруктозу, а лактаза - лактозу до глюкозы и галактозы. Клетчатка (целлюлоза) из-за отсутствия целлюлазы в животном организме не разлагается ферментами пищеварительных соков. Из кишечника в кровь всасываются только моносахариды. Скорость всасывания у разных моносахаридов различна. Полагают, что они всасываются в виде моносфорных эфиров, что дает возможность взаимопревращению в стенке кишечника гексоз, в частности, превращению фруктозы и галактозы в глюкозу. Моносахариды с током крови по системе воротной вены попадают в печень. В печени часть глюкозы превращается в гликоген. Печень способна как синтезировать гликоген, так и расщеплять его с образованием глюкозы.
ГЛИКОЛИЗ анаэробное (без участия О2) негидролитич. расщепление углеводов в цитоплазме под действием ферментов, сопровождающееся синтезом АТФ и заканчивающееся образованием молочной к-ты. Гликолиз одной молекулы глюкозы м. б. выражен след. ур-нием: C6H12O6+2H3PO4+2АДФ=2СН3СН(ОН)СООН+2АТФ – 135 кДж. Субстратами гликолиза кроме глюкозы м. б. другие моносахариды, а также полисахариды. У животных и человека в регуляции гликолиза принимают участие также гормоны. Гликолиз-простейшая форма биол. механизма аккумулирования энергии углеводов в АТФ. Считают, что он возник в период, когда в атмосфере Земли не было О2. При энергетически более выгодном аэробном окислении из одной молекулы глюкозы образуется 38 молекул АТФ. БРОЖЕНИЕ. Процессы анаэробного распада глюкозы, вызываемые различными микроорганизмами, известны под названием брожений. Наиболее распространенными видами брожений являются: 1. Спиртовое брожение. Его суммарное уравнение: С6Н120 ® 2 С2Н5ОН+2 СО2, 2.Молочнокислое брожение идет по уравнению:СбН12Об ® 2 СН3СНОН—СООН 3. Маслянокислое брожение:СбН120б —® СН3—СН2—СН2—СООН+2СО2+2Н2. Молочнокислое брожение играет большую роль при производстве молочнокислых продуктов (кефира, простокваши и др.), при квашении капусты, огурцов, при силосовании кормов. Маслянокислое брожение вызывается маслянокислыми бактериями, большинство из которых являются анаэробами. Образование масляной кислоты также идет через пировиноградную кислоту
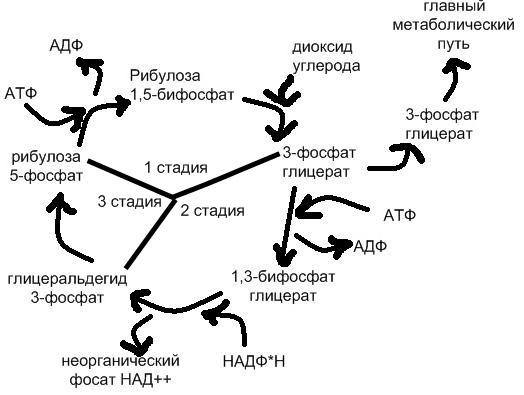 31.Пентозомонофосфатный путь. Цикл Кальвина.
31.Пентозомонофосфатный путь. Цикл Кальвина.
В цикл вовлекаются АТФ и НАДФ·Н, образованные в ЭТЦ фотосинтеза, углекислый газ и вода; основным продуктом является глицеральдегид-3-фосфат. Поскольку АТФ и НАДФ·Н могут образовываться в разных метаболических путях, цикл не следует рассматривать строго привязанным к световой фазе фотосинтеза. Общий баланс реакций цикла можно представить уравнением: 3 CO2 + 6 НАДФ·Н + 5 H2O + 9 АТФ → C3H5O3-PO3 + 3 H+ + 6 НАДФ+ + 9 АДФ + 8 Фн. Две молекулы глицеральдегид-3-фосфата используются для синтеза глюкозы. Цикл состоит из трёх стадий (карбоксилирование, восстановление, регенерация): на первой под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа происходит присоединение CO2 к рибулозо-1,5-дифосфату и расщепление полученной гексозы на две молекулы 3-фосфоглицериновой кислоты (3-ФГК). На второй 3-ФГК восстанавливается до глицеральдегид-3-фосфата (фосфоглицеральдегида, ФГА), часть молекул которого выходит из цикла для синтеза глюкозы, а другая часть используется в третьей стадии для регенерации рибулозо-1,5-дифосфата. Карбоксилирование. Карбоксилирование рибулозо-1,5-бисфосфата (5-углеродное соединение) осуществляется в несколько стадий. На первой кетонная группа рибулозы восстанавливается до спиртовой, между 2 и 3 атомами углерода устанавливается двойная связь. Полученное соединение нестабильно и именно оно карбоксилируется с образованием 2-карбокси-3-кето-D-арабитол-1,5-бисфосфата. Его структурный аналог 2-карбокси-D-арабитол-1,5-бисфосфат ингибирует весь процесс. Новое, уже 6-углеродное соединение, также нестабильно и распадается на две молекулы 3-фосфоглицериновой кислоты (3-фосфоглицерат, 3-ФГА). Восстановление. Восстановление 3-фосфоглицериновой кислоты (3-ФГА) происходит в две реакции. Сначала каждая 3-ФГА с помощью 3-фосфоглицераткиназы и с затратой одной АТФ фосфорилируется, образуя 1,3-бисфосфоглицериновая кислота (глицерат-1,3-бисфосфат). Затем под действием глицеральдегид-1,3-фосфатдегидрогеназы бисфосфоглицериновая кислота восстанавливается НАД(Ф)·H (у растений и цианобактерий; у пурпурных и зелёных бактерий восстановителем является НАД·H) параллельно с отщеплением одного остатка фосфорной кислоты. Образуется глицеральдегид-3-фосфат (фосфоглицеральдегид, ФГА, триозофосфат). Обе реакции обратимы. Регенерация. На последней стадии 5 молекул глицеральдегид-3-фосфатов превращаются в три молекулы рибулозо-1,5-бисфосфата. Вначале под действием трифосфат-изомеразы глицеральдегид-3-фосфат изомеризуется в дигидроксиацетон-фосфат. Фруктозабисфосфат-альдолаза объединяет их в фруктозо-6-фосфат с отщеплением остатка фосфорной кислоты. Затем следует ряд реакций перестройки углеродных скелетов и образуется рибулозо-5-фосфат. Он фосфорилируется фосфорибулокиназой и рибулозо-1,5-бисфосфат регенерируется.
32.Глюконеогенез, синтез моносахаридов из неуглеводных предшественников, происходящий в живых клетках под действием ферментов. Глюконеогенез осуществляется в направлении, обратном гликолизу. Большинство стадий этих двух процессов совпадают и катализируются одинаковыми ферментами.
Первая и вторая стадии этого процесса протекают в митохондриях. Образовавшаяся яблочная к-та способна проникать через мембрану митохондрий в цитоплазму и участвовать в дальнейших превращениях. У растений и бактерий обнаружены ферменты, осуществляющие синтез фосфоенолпиро-виноградной к-ты без промежут. стадий, а у нек-рых животных он протекает полностью в митохондриях, откуда эта к-та поступает в цитоплазму для участия в дальнейших р-циях глюконеогенеза. В цитоплазме может осуществляться также восстановительное карбоксилирование пировиноградной к-ты с образованием яблочной. Фруктозо-6-фосфат образуется в результате необратимого гидролиза фруктозо-1,6-дифосфата. Глюкозо-6-фосфат дефосфорилируется с образованием глюкозы или превращ. в глюкозо-1-фосфат-ключевое промежут. соед. в синтезе углеводов. Синтез одной молекулы глюкозы м. б. выражен суммарным ур-нием: 2СН3С(O)СООН + 2НАДН + 4АТФ + 2ГТФ -> -> С6Н12О6 + 2НАД + 4АДФ + 2ГДФ + 6Н3РО4 Осн. пункты контроля глюконеогенеза-регуляция синтезов фосфоенол-пировиноградной к-ты и глюкозо-6-фосфата. Первая р-ция катализируется пируваткарбоксилазой (активируется ацетилированным коферментом А), вторая - фруктозо-бис-фосфатазой (ингибируется аденозинмонофосфатом и активируется АТФ). Регуляция глюконеогенеза в организме человека и животных осуществляется также гормонами, напр. инсулин тормозит синтез ферментов глюконеогенеза, катехоламины, глюкагон и адренокортикотропин стимулируют глюконеогенез в печени, а паратиреоидный гормон-в почках.
|
|
|
|
|
Дата добавления: 2015-04-23; Просмотров: 2154; Нарушение авторских прав?; Мы поможем в написании вашей работы!