КАТЕГОРИИ:
Архитектура-(3434)Астрономия-(809)Биология-(7483)Биотехнологии-(1457)Военное дело-(14632)Высокие технологии-(1363)География-(913)Геология-(1438)Государство-(451)Демография-(1065)Дом-(47672)Журналистика и СМИ-(912)Изобретательство-(14524)Иностранные языки-(4268)Информатика-(17799)Искусство-(1338)История-(13644)Компьютеры-(11121)Косметика-(55)Кулинария-(373)Культура-(8427)Лингвистика-(374)Литература-(1642)Маркетинг-(23702)Математика-(16968)Машиностроение-(1700)Медицина-(12668)Менеджмент-(24684)Механика-(15423)Науковедение-(506)Образование-(11852)Охрана труда-(3308)Педагогика-(5571)Полиграфия-(1312)Политика-(7869)Право-(5454)Приборостроение-(1369)Программирование-(2801)Производство-(97182)Промышленность-(8706)Психология-(18388)Религия-(3217)Связь-(10668)Сельское хозяйство-(299)Социология-(6455)Спорт-(42831)Строительство-(4793)Торговля-(5050)Транспорт-(2929)Туризм-(1568)Физика-(3942)Философия-(17015)Финансы-(26596)Химия-(22929)Экология-(12095)Экономика-(9961)Электроника-(8441)Электротехника-(4623)Энергетика-(12629)Юриспруденция-(1492)Ядерная техника-(1748)
Вторичная структура
|
|
|
|
Структура полинуклеотидной цепи
Структура нуклеиновых кислот
Нуклеиновые кислоты представляют линейные полимеры нуклеозидмонофосфатов, то есть полинуклеотиды. Нуклеотиды построены из трех компонентов: пиримидинового или пуринового основания, пентозы и фосфорной кислоты. Нуклеотиды связаны между собой в цепь фосфодиэфирной связью. Она образуется за счет этерификации ОН — группы С—З- пентозы одного нуклеотида и ОН — группы фосфатного остатка другого нуклеотида. В результате один из концов полинуклеотидной цепи заканчивается свободным фосфатом (Р—конец или5-—конец). На другом конце цепи имеется неэтерифицированная ОН — группа у С—З- пентозы (З- — конец).
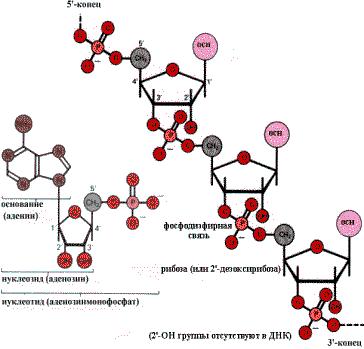
Типичный нуклеотид (АМР)
Первичная структура нуклеиновых кислот определяется как последовательность нуклеотидных остатков в полимерной цепи. Многообразие молекул DNA и RNA объясняется их первичной структурой. Как многие другие биополимеры, нуклеиновые кислоты имеют ещё и вторичную структуру, под которой понимают их пространственную организацию.
DNA Молекула DNA представляет собой правозакрученную спираль, состоящую из двух полинуклеотидных цепей с антипараллельным ходом. Это означает, что 3-—концу одной цепи соответствует 5-—конец другой цепи и наоборот.
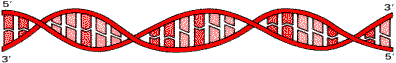
Структура двойной спирали DNA
Остатки оснований направлены внутрь спирали. На один виток спирали приходится 10 пар оснований. Цепи DNA не идентичны, так как нуклеотидный состав их различен, однако первичная структура одной цепи предопределяет нуклеотидную последовательность другой цепи, то есть они комплементарны друг другу. Это связано с существованием комплементарных пар оснований.
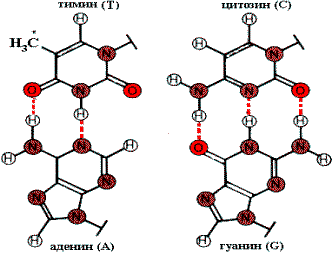
Комплементарные пары оснований. *СН3—группа в тимине (Т) замещается на H в урациле (U).
Физико—химическую основу комплементарности составляют водородные связи, которые могут образоваться только между аденином одной цепи и тимином другой, противоположно направленной цепи (пара А—Т), и аналогично между гуанином и цитозином (пара G—C). Вторичная структура RNA несколько иная. Молекула RNA состоит из одной полинуклеотидной цепи. Отдельные участки этой цепи (до 20—30 нуклеотидных пар) могут быть комплементарны между собой и образовывают спиральную структуру за счет связей между аденином и урацилом (пара A—U) и гуанином и цитозином (пара G—C). Между спирализованными участками располагаются одноцепочечные петли. Существует несколько разновидностей RNA: матричная (mRNA), транспортная (tRNA), рибосомная (rRNA). На рисунке приведена структура tRNA, у которой спирализованные участки определяют специфическую пространственную конформацию: фигуру Lклеверного листа¦.
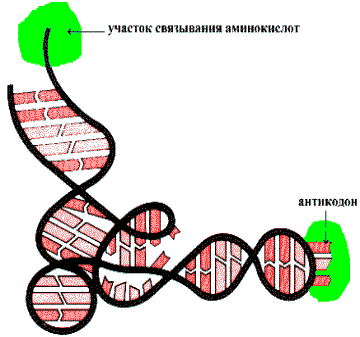
Трехмерная пространственная структура транспортной RNA
Анимация трехмерной пространственной структуры транспортной RNA
tRNA имеет на 3-—конце ССА для связывания аминокислоты, а в средней части молекулы — антикодоновый участок — последовательность нуклеотидов, обеспечивающую взаимодействие tRNA с кодоном mRNA.
Матричный cинтез. 1. Полимеризация и поликонденсация, при которых строение образующегося полимера и (или) кинетика процесса определяются другими макромолекулами (матрицами), находящимися в непосредственном контакте с молекулами одного или неск. мономеров и растущими цепями. Пример матричного синтеза в живой природе - синтез нуклеиновых кислот и белков. в котором роль матрицы играют ДНК и РНК, а состав и порядок чередования звеньев в растущей (дочерней) цепи однозначно определяются составом и структурой матрицы.
Термин "матричный синтез " обычно используют при описании синтеза нуклеиновых кислот и белков. а при рассмотрении способов получения др. полимеров пользуются такими терминами, как матричные полиреакции, полимеризация, поликонденсация. Такой матричный синтез реализуется при условии хим. и стерич. соответствия (комплементарности) мономеров и растущей цепи, с одной стороны, и матрицы - с другой; при этом элементарные акты осуществляются между мономерами и растущими макромолекулами (а также олигомерами - при матричной поликонденсации), связанными с матрицей.
Обычно мономеры и олигомеры обратимо связываются с матрицей достаточно слабыми межмолекулярными взаимодействиями - электростатическими, донорно-акцепторным и т.д. Дочерние цепи практически необратимо ассоциируют с матрицей ("узнают" матрицу) только после того, как достигнут некоторой определенной длины, зависящей от энергии взаимодействия между звеньями матрицы и дочерней цепи. "Узнавание" матрицы растущей цепью - необходимая стадия матричный синтез; дочерние цепи практически всегда содержат фрагмент или фрагменты, образовавшиеся по "обычному" механизму, т. е. без влияния матрицы. Скорость матричный синтез может быть выше, ниже или равна скорости процесса в отсутствие матрицы (кинетический матричный эффект).
Структурный матричный эффект проявляется в способности матрицы влиять на длину и химическое строение дочерних цепей (в том числе их стерическую структуру), а если в матричном синтезе участвуют два или более мономера - то также на состав сополимера и способ чередования звеньев.
Методом матричного синтеза получают полимер-полимерные комплексы, обладающие более упорядоченной структурой, чем поликомплексы, синтезируемые простым смешением растворов полимеров, а также поликомплексы, которые нельзя получить из готовых полимеров вследствие нерастворимости одного из них. Матричный синтез - перспективный метод получения новых полимерных материалов.
Термин "матричный синтез " обычно используют при описании синтеза нуклеиновых кислот и белков. а при рассмотрении способов получения др. полимеров пользуются такими терминами, как матричные полиреакции, полимеризация, поликонденсация. Лит.: Кабанов В. А., Паписов И. М., "Высокомолекулярные соединения", сер. А, 1979, т. 21, № 2, с. 243-81; Картина О. В. [и др.], "ДАН СССР", 1984, т. 275, №3, с. 657-60; Литманович А. А., Марков С. В., Паписов И. М., "Высокомолекулярные соединения", сер. А, 1986, т. 28, №6, с. 1271-78; Ferguson J., Al-Alawi S., Graumayen R., "European Polymer Journall", 1983, v. 19, № 6, p. 475-80; Polоwinski S., "J. Polymer. Sci.", Polimer Chemistry Edition, 1984, v. 22, № 11, p. 2887-94. И. М. Паписов.
2. Химические реакции, в которых строение образующегося мономолекулярного органического соединения и (или) кинетика процесса определяется атомом металла (так называемый темплатный синтез).
Атом металла может входить в состав соли или комплексного соединения и выполнять в матричном синтезе различные функции. Он координирует молекулы и тем самым ориентирует их реагирующие фрагменты (так называемый кинетический эффект в матричном синтезе); в этом случае образование целевого продукта без участия в реакции атома металла вообще не происходит. Атом металла может связывать в комплекс только один из конечных продуктов, которые образуются в равновесной реакции (так называемый термодинамический эффект в матричном синтезе); образование целевого продукта может происходить и в отсутствие металла, однако под влиянием последнего выход реакции существенно возрастает. Часто оба эти механизма проявляются одновременно. Известны случаи, когда равновесная реакция осуществляется на стадии образования промежуточного продукта. Последний фиксируется в виде металлокомплекса, и дальнейшее превращение идет специфическим образом (так называемый равновесный эффект в матричном синтезе). Возможны и другие механизмы матричного синтеза.
Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул мРНК и тРНК.
[править] Введение
Биосинтез белка можно разделить на стадии транскрипции, процессинга и трансляции. Во время транскрипции происходит считывание генетической информации, зашифрованной в молекулах ДНК, и запись этой информации в молекулы иРНК. В ходе ряда последовательных стадий процессинга из мРНК удаляются некоторые фрагменты, ненужные в последующих стадиях, и происходит редактирование нуклеотидных последовательностей. После транспортировки кода из ядра к рибосомам происходит собственно синтез белковых молекул, путём присоединения отдельных аминокислотных остатков к растущей полипептидной цепи.
[править] Процессинг РНК
Основная статья: Процессинг РНК
Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки. К 5΄-концу присоединяется кэп, а к 3΄-концу поли-А хвост, который увеличивает длительность жизни иРНК. С появлением процессинга в эукариотической клетке стало возможно комбинирование экзонов гена для получения большего разнообразия белков, кодируемым единой последовательностью нуклеотидов ДНК, — альтернативный сплайсинг.
[править] Трансляция
Основная статья: Трансляция (биология)
Трансляция заключается в синтезе полипептидной цепи в соответствии с информацией, закодированной в матричной РНК. Аминокислотная последовательность выстраивается при помощи транспортных РНК, которые образуют с аминокислотами комплексы — аминоацил-тРНК. Каждой аминокислоте соответствует своя тРНК, имеющая соответствующий антикодон, «подходящий» к кодону мРНК. Во время трансляции рибосома движется вдоль мРНК, по мере этого наращивается полипептидная цепь. Энергией биосинтез белка обеспечивается за счёт АТФ.
Готовая белковая молекула затем отщепляется от рибосомы и транспортируется в нужное место клетки. Для достижения своего активного состояния некоторые белки требуют дополнительной посттрансляционной модификации.
|
|
|
|
|
Дата добавления: 2015-04-23; Просмотров: 769; Нарушение авторских прав?; Мы поможем в написании вашей работы!